Возрастные изменения структуры и функции скелетных мышц
Описаны возрастные изменения структуры и функции скелетных мышц. Причину возрастной атрофии (саркопении) скелетных мышц авторы видят в уменьшении количества ДЕ. Приведены данные о возрастной потере количества мышечных волокон и ДЕ в латеральной широкой мышце бедра.

Faulkner, J.A. Age-related changes in the structure and function of skeletal muscles / J.A. Faulkner, L. M. Larkin, D.R. Claflin, S. V. Brooks // Proceedings of the Australian Physiological Society, 2007. – 38. – p. 69-75.
ДЖ. А. ФОЛКНЕР С СОАВТ.
ВОЗРАСТНЫЕ ИЗМЕНЕНИЯ СТРУКТУРЫ И ФУНКЦИИ СКЕЛЕТНЫХ МЫШЦ
АННОТАЦИЯ
- Для животных всех возрастов в период сокращения скелетных мышц баланс между силой, развиваемой мышцами, и внешней нагрузкой определяет, будет ли мышца укорачиваться, оставаться при постоянной длине (изометрической) или удлиняться. При максимальной активации сила, развиваемая в течение укорочения, минимальна, средняя (промежуточное значение) — когда длина мышцы фиксирована, и максимальная во время удлинения. Во время работы в уступающем режиме, когда усилие высоко, мышцы могут быть повреждены из-за сокращения.
- Слабость, «потеря веса» и остановка в физическом развитии» наиболее часто наблюдаются у пожилых, физически неактивных людей. Слабый человек определяется как человек с маленьким ростом и с мышцами, которые атрофировались, слабый и легко устающий. Состояние «отсутствия прибавки в весе и остановки в физическом развитии» олицетворяет отсутствие ответа на хорошо разработанные программы питания и физической активности.
- При старении атрофия скелетных мышц человека является неизбежной. Постепенная потеря мышечных волокон начинается в среднем в возрасте 50 лет и продолжается вплоть до 80 лет. Исследования показывают, что приблизительно 50% волокон теряются из мышц конечностей. Для людей и для крыс, наблюдалось, что сроки и величина потерь двигательных единиц (ДЕ) аналогичны. Это означает, что механизм ответственный за потерю мышечных волокон и ДЕ у них идентичен. Степень атрофии мышечных волокон, во многом зависит от привычного уровня физической активности индивидуума.
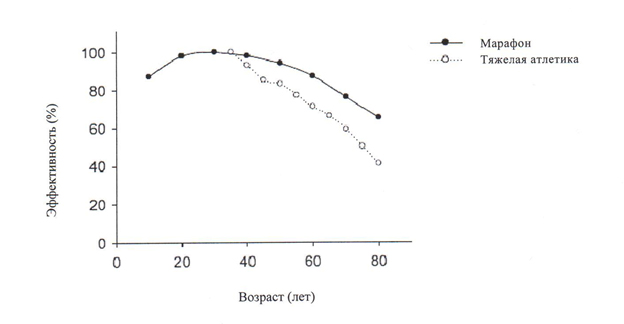
- Спортсмены высокой квалификации поддерживают высокий уровень физической формы на протяжении всего периода их жизни. Даже среди профессиональных спортсменов, эффективность марафонцев и тяжелоатлетов снижается после 40 лет, максимальный уровень производительности снижается приблизительно на 50% к 80 годам. Успех у профессиональных спортсменов и у малоподвижных пожилых людей, совершающих хорошо продуманные, тщательно управляемые тренировочные программы обеспечивают впечатляющее доказательство того, что возрастная атрофия, слабость и утомляемость могут быть замедлены, но не остановлены.
ТИПЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ
У живых организмов всех возрастов и всех видов скелетные мышцы при активации могут выполнять три типа сокращений (1). Если внешняя нагрузка на мышцу меньше напряжения в мышце, то волокна, и, следовательно, мышца будет укорачиваться. И наоборот, если нагрузка превышает напряжение, то мышечные волокна растянутся. А если нагрузка и сила равны или если нагрузка статическая, то мышечные волокна останутся “фиксированными” в той же длине, то есть проявится изометрический тип сокращения (Рисунок 1). По сравнению с силой, развиваемой максимально активированными скелетными мышцами в момент изометрического сокращения, максимальная сила, развиваемая в момент растяжения – в 1,5-2 раза больше, в то время как сила, развиваемая при сокращении всегда меньше максимальной изометрической силы (Рисунок 1). Мышца выполняет «работу» только в момент сокращения, и величина совершаемой работы – это функция силы, созданная мышцей и величиной сокращения (Рисунок 1). При повторных изометрических сокращениях или сокращениях в преодолевающем режиме, или растяжении без активации, сила может уменьшаться из-за утомления, но восстановление силы до контрольных значений происходит достаточно быстро и в полной мере, никаких признаков травм не наблюдается (2,3).
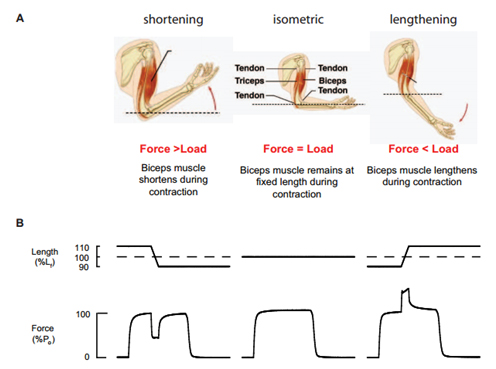
В противоположность этому, повреждение мышцы при сокращении происходит только при ее работе в уступающем режиме (2,3). Faulkner & Brooks (4) выяснили, что травма провоцирует механическое разрушение отдельных саркомеров с последующим блокированием очага травмы (5). Более серьезное дальнейшее повреждение проявляется спустя несколько дней из-за появления в мышцах, воспалительных клеток и образования свободных радикалов (4,6,7). Травмы скелетной мышцы часто связаны либо с одним сильным растяжением максимально активированной мышцы, например, во время падения, или сразу нескольких небольших растяжений сильно активизированной мышцы, как, например, мышцы бедра и разгибатели колена во время марафонского бега (4). Полное восстановление после травмы, вызванной растяжением, требуется несколько недель, а если растяжения тяжелые, то даже больше (4,6, 7). У старых животных работа в эксцентрическом режиме ведет к большему количеству повреждения, при этом после тяжелого повреждения восстановление может быть неполным (4). Неполное восстановление повреждения, вызванного сокращением, может привести к необратимой потере мышечной массы и силы (6,7).
СЛАБОСТЬ И НЕСПОСОБНОСТЬ РАЗВИВАТЬСЯ
Борт (8) идентифицировал опорно-двигательный аппарат как «путь вступления для слабости». Следовательно, структурно-функциональные факторы, ответственные за физический аспект слабости являются наиболее очевидными и наиболее эффективно исследованы непосредственно в скелетных мышцах человека (9-12). В этих исследованиях однозначно установлено 30% — 50% уменьшение массы скелетных мышц у мужчин и женщин в возрасте от 40 до 80 лет (12-15). Для большинства пожилых людей, снижение мышечной массы (12, 15) сопровождается, по крайней мере, равным, а обычно даже большим снижением силы (12, 16, 17) и мощности (18), а также увеличением удельной силы (силы на единицу поперечного сечения площади мышцы) (19) и утомляемостью (20, 21).
Общим результатом такого воздействия являются возрастные изменения в опорно-двигательном аппарате, оказывающие значительное влияние на повседневные действия пожилых людей. Это воздействие еще более существенно для «слабых» пожилых людей. Состояние «физической слабости» у людей ассоциируется с человеком маленького роста, у которого представлены серьезные нарушения мышечной силы, подвижности, равновесия и выносливости из-за слабой и быстро утомляющейся мускулатуры (22, 23). Состояние слабости только изредка наблюдается у молодых людей или даже людей средних лет, но чрезвычайно распространено среди пожилых людей. Как принято считать, причины «слабости» – это генетика, болезнь, повреждения, образ жизни и старение (8). Хотя состояние «слабости» связано со многими болезнями, для здоровых пожилых людей это состояние, как оказывается связано, прежде всего, с образом жизни (8). Кроме того, у многих слабых пожилых людей наблюдается феномен «остановки в весе», при этом «слабый пожилой человек» не поддается лечению по специально разработанным программам для улучшения физиологического состояния и реабилитации (24).
Состояния «слабости» и «остановки в весе» составляют два наиболее заметных проявления у малоподвижных пожилых людей. В любом возрасте слабость может возникнуть из-за метаболических болезней или мышечной атрофии, или длительного неактивного образа жизни (8), но для молодых людей такое состояние обычно обратимо. Повышенная восприимчивость к размеру повреждений, вызванных сокращением активности, их масштабу, составляют дополнительный фактор риска для пожилых людей (6,7). Поэтому «слабость» может привести к сниженной подвижности и потере равновесия (25), более высокому риску падения (26,27), повышенному уровню и серьезности повреждений, вызванных мышечным сокращением (4). Состояние слабости неизменно сопровождается потерей физической формы, здоровья и качества жизни (28). Сопутствующая состоянию «слабости» «остановка в весе» отражает неспособность слабого пожилого человека высвободиться из нисходящей спирали всё большей и большей слабости, даже когда такая возможность имеется.
МЕХАНИЗМЫ, ОТВЕТСТВЕННЫЕ ЗА АТРОФИЮ СКЕЛЕТНЫХ МЫШЦ
Мышечная масса является функцией от среднего объема всех мышечных волокон (длина мышечных волокон, умноженная на площадь поперечного сечения (ППС) и числа волокон, имеющихся в данной скелетной мышце. В зрелом возрасте длина мышечных волокон изменяется только в сочетании со значительной гипертрофией или атрофией. (29). Более того, несмотря на то, что возможно расщепление мышечных волокон (30), их количество не увеличивается (31). Вследствие этого, для взрослых людей изменение массы данной мышцы происходит прежде всего из-за изменения в поперечного сечении мышечных волокон или потери определенного количества волокон. У большинства людей оба фактора способствуют уменьшению мышечной массы по мере старения, но доля снижения у каждого индивида зависит от наследственности так же, как и от ряда неизвестных факторов в дополнение к привычным уровням физической активности (16). Для многих атрофия скелетных мышц первично замечается после 40 лет и практически у всех после 50 лет (15). Возникающая потеря мышечной массы до 50 лет напрямую связана с уменьшением площади поперечного сечения отдельных волокон из-за сидячего образа жизни, поскольку не наблюдается различий в количестве волокон от 20 вплоть до 50 лет (Рисунок 2а). Между 50 и 80 годами жизни число волокон в латеральной широкой мышце бедра у мужчин уменьшается с 600000 волокон до 323000 волокон (Рисунок 2а). Хотя сопоставимое исследование для женщин не проводилось, возрастные изменения в мышечной массе предполагают аналогичные изменения (16,18). Неизбежно теряется часть мышечных волокон 2 типа (11,15), но потеря мышечной массы в значительной степени зависит от регулярности и интенсивности физической активности, в которую вовлечены пожилые люди. Физическая активность непосредственно воздействует на ППС оставшихся мышечных волокон, и вместо атрофии, волокна могут сохранять свою ППС (32,33), или даже гипертрофироваться (10,11, 34). Тенденция такова — ППС быстрых мышечных волокон 2 типа с возрастом уменьшается, в то время как медленные волокна 1 типа сохраняют свою ППС, даже у пожилых индивидов (11, 15).
Крупные мышцы бедра человека состоят из сотен тысяч волокон (15). Несмотря на значительные проблемы в получении образцов из мышц с помощью игольчатой биопсии и влияния площади поперечного сечения на измерения, полученные данные оказались достоверными (32,33,35). Исследования мужчин и женщин, как молодых, так и старшей возрастной группы, проводившиеся с учетом тщательно разработанных программ тяжелой атлетики и при условии единых показателей структуры и функционирования проницаемых тканей, продемонстрировали гипертрофию в мышечных волокнах 1 и 2 типа (32, 35). В остальных случаях изменений в площади поперечного сечения не было (33). Расхождения в этих данных вероятно связаны с различиями в первоначальном уровне физической подготовки исследуемых, в интенсивности программы тренировок, а также в том, что при биопсии изучаются только несколько сотен мышечных волокон из крупных мышц бедра (32-35).
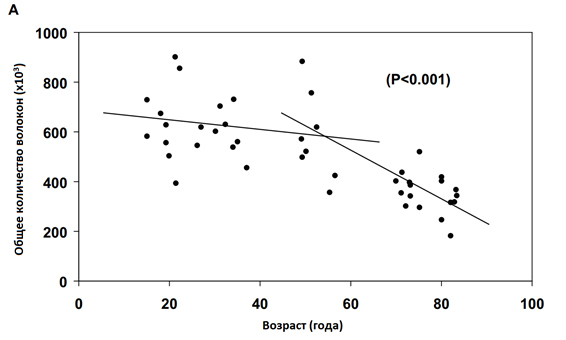
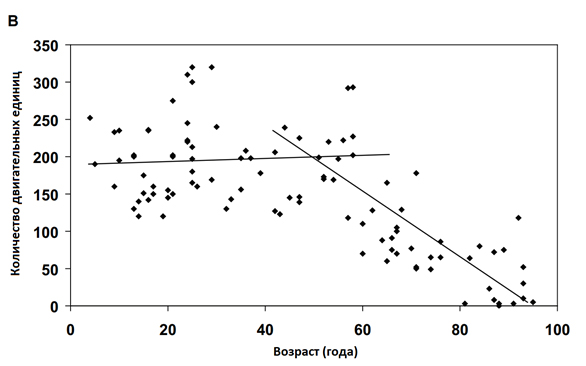
МЕХАНИЗМЫ, ОТВЕТСТВЕННЫЕ ЗА ПОТЕРЮ МЫШЕЧНЫХ ВОЛОКОН
Уменьшение мышечной массы и количества мышечных волокон наблюдается у каждого вида, у которого были изучены возрастные изменения: людей (11, 15), крыс (36-39) и мышей (40). Хотя происхождение многих механизмов, лежащих в основе возрастных изменений в скелетных мышцах остается неизвестным, однако четко установлен вклад денервации в потерю мышечных волокон (41), двигательных единиц (42-44), и их ремоделирование (45). Потеря двигательных единиц (ДЕ) стала основополагающим фактором в возрастных изменениях скелетных мышц и являлась причиной потери мышечных волокон (Рисунок 2В). Феномен потери ДЕ наблюдается как в мышцах крыс (43, 45-47), так и в мышцах человека (42, 48, 49). Хотя, у людей количество ДЕ измерялось в мышцах ладони (48,49) и пальцев (42) с помощью косвенных методов, в то время как у крыс подсчитывали количество волокон напрямую в крупных мышцах бедра (15).
В то время как наблюдения за возрастными изменениями, связанными со скелетными мышцами человека являются наиболее актуальными (15,42, 48-50), исследования основополагающих механизмов связанных с возрастными изменениями в скелетных мышцах мелких, непродолжительно живущих видов млекопитающих, таких как мыши (20,40) и крысы (45), обеспечивают много преимуществ. Преимущества модели грызунов очевидно подходят для менее точных переменных, а для организма человека возможно применение косвенных методов исследования. Такие переменные включают в себя: измерение всей мышечной массы (40) количества мышечных волокон (36, 49), абсолютную и удельную силу всех мышц (40), абсолютную мощность и нормированное напряжение всех мышц (20), а также количество ДЕ, коэффициент иннервации и ремоделирование (43, 45-47). Учитывая короткий период жизни крыс, от двух до трех лет, сроки начала, скорость и величина потери ДЕ крыс (43, 47) находятся в хорошем соответствии с потерями ДЕ у человека (Рисунок 2). Кроме того, количество аксонов ДЕ, иннервирующих скелетные мышцы, уменьшается как у старых крыс (41, 46, 51), так и у пожилых людей (52). Сокращение количества мотонейронов и количества ДЕ в старости у человека, сравнивалось со зрелыми крысами, и их величины оказались одинаковыми (46). Несмотря на эту взаимосвязь, никакой причинно-следственной связи между потерей числа аксонов в мотонейронах, и потерей ДЕ и мышечных волокон не было установлено, как у крыс, так и у человека (53).В дополнение к потере ДЕ в качестве существенной причины возникновения мышечной атрофии, возможна денервационная атрофия отдельных мышечных волокон независимо от потери ДЕ (54, 55). У крыс потеря ДЕ происходит исключительно в пулах быстрых, мощных ДЕ (45). Потеря быстрых ДЕ оставляет несколько быстрых волокон в денервируемой мышце. Некоторые из этих волокон включаются в оставшиеся медленные ДЕ с помощью прорастания аксонов (55). Результатом является увеличение числа мышечных волокон в оставшихся ДЕ, т.е. увеличение иннервационного коэффициента (45). Волокна, которые не стали повторно иннервироваться, подвергаются денервационной атрофии и в конце концов исчезают. В то время как потеря ДЕ имеет примерное соответствие между крысами (43,47) и людьми (42,48), количество мышечных волокон, исчезнувших при старении, не имеет такого соответствия между этими двумя видами. Для больших мышц конечностей человека, при достижении 80-летнего возраста происходит 50%-ое уменьшение числа волокон, в то время как мышцы задних конечностей крыс показывают только от 5-10% потери волокон за сопоставимый период их жизни (36,39). Подобная потеря ДЕ между видами в сочетании с гораздо большей потерей мышечных волокон от человека по сравнению с мышцами грызунов, свидетельствует о том, что в мелких мышцах крыс, процесс повторной иннервации является гораздо более эффективным, чем в больших мышцах человека, хотя эта гипотеза не была экспериментально проверена.
ПРИВЫЧНЫЙ УРОВЕНЬ И ТИП ФИЗИЧЕСКОЙ АКТИВНОСТИ
В прошедшей четверти двадцатого века существовали различия во мнениях по поводу развития неизменных возрастных изменений в структуре и функциях скелетных мышц (15, 42, 48) у людей с малоподвижным образом жизни и низкой физической активностью (17, 50). Среди взрослых мужчин и женщин, выделяют очень физически активных в любой возрастной группе (спортсмены высокой квалификации). На протяжении их жизненного пути, за исключением травм или болезней, эти спортсмены поддерживают высокий уровень физической подготовки и конкурируют в виде спорта или специальных индивидуальных программах по бегу, метанию или в тяжелой атлетике. Даже у этих мужчин и женщин производительность снижается к 40 годам. К 80 годам, снижение максимальной производительности составляет от 35% до 65% для различных видов спорт (рисунок 3). Вариативность между соревнованиями во многом зависит от того, включает ли соревнование перемещение массы тела, как в беге, или перемещение фиксированной массы, как в тяжелой атлетике, толкании ядра или метании диска. Результаты во всех дисциплинах и во всех возрастных группах были улучшены за счет появления «плиометрических тренировок» (56). «Плиометрические тренировки» включают в себя энергичные движения, которые связаны с каждым из трех типов сокращений: укорочение (концентрический режим), изометрический режим и растяжение (эксцентрический режим), как описано в первом разделе. Значительное увеличение мышечной массы, силы, мощности и выносливости были также достигнуты ранее у мужчин и женщин старше 75 лет ведущих малоподвижный образ жизни с помощью участия в комбинированных программах, которые включают: 1. Тренировки три раза в неделю в различные дни; 2. Сокращение мышц с участием каждого из трех типов сокращений; 3. Непрерывный прирост нагрузки на основе увеличения силы; и 4. Длительность тренировок 12 недель и более (32, 33, 35, 56, 57). Улучшения в таких программах тренировок значительно возросли за счет включения эксцентрических сокращений при выполнении одного движения с весом большим 80% от максимальной силы, но такие программы требуется выполнять под руководством профессиональных инструкторов и осуществлять с большой осторожностью, чтобы не вызвать у участников «повреждения мышц, вызванного сокращением» (6, 7, 56, 57). Успех спортсменов высокой квалификации и у пожилых людей, ранее ведущих сидячий образ жизни, которые взяли на себя обязательство выполнять хорошо разработанные, тщательно управляемые тренировочные программы, показывает, что атрофию мышц, слабость и утомляемость, которые обычно ассоциируются с преклонным возрастом, можно замедлить. Хотя потеря в количестве волокон в мышцах неизменно появляется, величина потери мышечной массы может быть в какой-то степени смягчена за счет гипертрофии волокон, которые остаются (32, 35).
Таким образом, ясно, что необратимые изменения в структуре и функционировании скелетных мышц, а также все более и более сидячий образ жизни способствуют увеличению физической слабости пожилых людей. Молодые люди и даже люди зрелого возраста способны без труда восстановиться после периодов слабости, добровольно или непреднамеренно вызванных физическим бездействием, травмой, голодом или болезнью (23,50). Несмотря на значительные достижения в качестве и научном прогрессе изучения программ по развитию физической формы для ослабленных пожилых людей (23,24,58), феномен «отставания в развитии» продолжает быть непостижимым аспектом даже для хорошо разработанных программ по созданию физической формы (8,58). В США стоимость физической слабости среди пожилых людей оценивается в миллионы долларов в год, и несмотря на это растет число ослабленных пожилых людей, возраст которых предположительно будет начинаться с 40 лет (59). Несмотря на огромный рост затрат и серьезность проблемы (59), было предложено не так много идей и сделаны лишь скромные успехи в создании успешных программ физического развития (23) для ослабленных пожилых людей и решения феномена «отставание в развитии» (8,58).
REFERENCES
- Faulkner JA. Terminology for contractions of muscles during shortening, while isometric, and during lengthening. J. Appl. Physiol. 2003; 95: 455-9.
- McCully KK, Faulkner JA. Injury to skeletal muscle fibers of mice following lengthening contractions. J. Appl. Physiol. 1985; 59: 119-26.
- McCully KK, Faulkner JA. Characteristics of lengthening contractions associated with injury to skeletal muscle fibers. J. Appl. Physiol. 1986; 61: 293-9.
- Faulkner JA, Brooks SV, Zerba E. Muscle atrophy and weakness with aging: contraction-induced injury as an underlying mechanism. J. Gerontol. 1995; 50A: 124-9.
- Rader EP, Song W, Van Remmen H, Richardson A, Faulkner JA. Raising the antioxidant levels within mouse muscle fibres does not affect contractioninduced injury. Exp. Physiol. 2006; 91: 781-9.
- Rader EP, Faulkner JA. Recovery from contractioninduced injury is impaired in weight-bearing muscles of old male mice. J. Appl. Physiol. 2006; 100: 656-61.
- Rader EP, Faulkner JA. Effect of aging on the recovery following contraction-induced injury in muscles of female mice. J. Appl. Physiol. 2006; 101: 887-92.
- Bortz WM. A conceptual framework of frailty: a review. J. Gerontol. A. Biol. Sci. Med. Sci. 2002; 57: M283-M288.
- Aniansson A, Hedberg M, Henning GB, Grimby G. Muscle morphology, enzymatic activity, and muscle strength in elderly men: a follow-up study. Muscle Nerve 1986; 9: 585-91.
- Grimby G, Saltin B. The ageing muscle. Clin. Physiol. 1983; 3: 209-18.
- Lexell J. Human aging, muscle mass, and fiber type composition. J. Gerontol. 1995; 50A: 11-6.
- Young A, Stokes M, Crowe M. The size and strength of the quadriceps muscles of old and young men. Clin. Physiol. 1985; 5: 145-54.
- Akima H, Kano Y, Enomoto Y, Ishizu M, Okada M, Oishi Y, Katsuta S, Kuno S. Muscle function in 164 men and women aged 20-84 yr. Med. Sci. Sports Exerc. 2001; 33: 220-6.
- Allen TH, Andersen EC, Langham WH. Total body potassium and gross body composition in relationship to age. J. Gerontol. 1960; 15: 348-57.
- Lexell J, Taylor CC, Sjostrom M. What is the cause of the ageing atrophy? Total number, size and proportion of different fiber types studied in whole vastus lateralis muscle from 15- to 83-year-old men. J. Neurol. Sci. 1988; 84: 275-94.
- Frontera WR, Hughes VA, Lutz KJ, Evans WJ. A cross-sectional study of muscle strength and mass in 45- to 78-yr-old men and women. J. Appl. Physiol. 1991; 71: 644-50.
- Goodpaster BH, Park SW, Harris TB, Kritchevsky SB, Nevitt M, Schwartz AV, Simonsick EM, Tylavsky FA, Visser M, Newman AB. The loss of skeletal muscle strength, mass, and quality in older adults: the health, aging and body composition study. J. Gerontol. A. Biol. Sci. Med. Sci. 2006; 61: 1059-64.
- Bassey EJ, Fiatarone MA, O’Neill EF, Kelly M, Evans WJ, Lipsitz LA. Leg extensor power and functional performance in very old men and women. Clin. Sci. (Lond) 1992; 82: 321-7.
- Brooks SV, Faulkner JA. Skeletal muscle weakness in old age: underlying mechanisms. Med. Sci. Sports Exerc. 1994; 26: 432-9.
- Brooks SV, Faulkner JA. Forces and powers of slow and fast skeletal muscles in mice during repeated contractions. J. Physiol. (Lond) 1991; 436: 701-10.
- Gandevia SC, Enoka RM, McComas AJ, Stuart DG, Thomas CK. Fatigue: neural and muscular mechanisms. Adv. Exp. Med. Biol. 1995; 384.
- Fried LP. Conference on the physiologic basis of frailty. Aging (Milano.) 1992; 4: 251-65.
- Hadley EC, Ory MG, Suzman R, Weindruch R, Fried L. Physical frailty: A treatable cause of dependence in old age. J. Gerontol. 1993; 48: 1-88.
- Verdery RB. Failure to thrive in the elderly. Clin. Geriatr. Med. 1995; 11: 653-9.
- Schultz AB. Muscle function and mobility biomechanics in the elderly: an overview of some recent research. J. Gerontol. 1995; 50A: 60-3.
- Lord SR, Sherrington C, Menz HB. Falls in Older People. Risk Factors and Strategies for Prevention, Cambridge University Press, Cambridge. 2001.
- Tinetti ME, Williams TF, Mayewski R. Fall risk index for elderly patients based on number of chronic disabilities. Am. J. Med. 1986; 80: 429-34.
- Baumgartner RN, Koehler KM, Gallagher D, Romero L, Heymsfield SB, Ross RR, Garry PJ, Lindeman RD. Epidemiology of sarcopenia among the elderly in New Mexico. Am. J. Epidemiol. 1998; 147: 755-63.
- Maxwell LC, Faulkner JA, Hyatt GJ. Estimation of number of fibers in guinea pig skeletal muscles. J. Appl. Physiol. 1974; 37: 259-64.
- Blaivas M, Carlson BM. Muscle fiber branching—difference between grafts in old and young rats. Mech. Ageing Dev. 1991; 60: 43-53.
- Gollnick PD, Timson BF, Moore RL, Riedy M. Muscular enlargement and number of fibers in skeletal muscles of rats. J. Appl. Physiol. 1981; 50: 936-43.
- Trappe S, Williamson D, Godard M, Porter D, Rowden G, Costill D. Effect of resistance training on single muscle fiber contractile function in older men. J. Appl. Physiol. 2000; 89: 143-52.
- Trappe S, Godard M, Gallagher P, Carroll C, Rowden G, Porter D. Resistance training improves single muscle fiber contractile function in older women. Am. J. Physiol. (Cell) 2001; 281: C398-C406.
- Lexell J, Taylor CC. Variability in muscle fibre areas in whole human quadriceps muscle: effects of increasing age. J. Anat. 1991; 174: 239-49.
- Widrick JJ, Stelzer JE, Shoepe TC, Garner DP. Functional properties of human muscle fibers after short-term resistance exercise training. Am. J. Physiol. (Regul. Integr. Comp. Physiol. ) 2002; 283: R408-R416.
- Daw CK, Starnes JW, White TP. Muscle atrophy and hypoplasia with aging: impact of training and food restriction. J. Appl. Physiol. 1988; 64: 2428-32.
- Eddinger TJ, Moss RL, Cassens RG. Fiber number and type composition in extensor digitorum longus, soleus, and diaphragm muscles with aging in Fisher 344 rats. J. Histochem. Cytochem. 1985; 33: 1033-41.
- Hooper AC. Length, diameter and number of ageing skeletal muscle fibres. Gerontology 1981; 27: 121-6.
- Larkin LM, Kuzon WM, Halter JB. Effects of age and nerve-repair grafts on reinnervation and fiber type distribution of rat medial gastrocnemius muscles. Mech. Ageing Dev. 2003; 124: 653-61.
- Brooks SV, Faulkner JA. Contractile properties of skeletal muscles from young, adult and aged mice. J. Physiol. (Lond) 1988; 404: 71-82.
- Ansved T, Larsson L. Quantitative and qualitative morphological properties of the soleus motor nerve and the L5 ventral root in young and old rats. Relation to the number of soleus muscle fibers. J. Neurol. Sci. 1990; 96: 269-82.
- Campbell MJ, McComas AJ, Petito F. Physiological changes in ageing muscles. J. Neurol. Neurosurg. Psychiatry 1973; 36: 174-82.
- Cederna PS, Asato H, Gu X, van der MJ, Kuzon WM, Jr., Carlson BM, Faulkner JA. Motor unit properties of nerve-intact extensor digitorum longus muscle grafts in young and old rats. J. Gerontol. A. Biol. Sci. Med. Sci. 2001; 56: B254-B258.
- McComas AJ, Fawcett PR, Campbell MJ, Sica RE. Electrophysiological estimation of the number of motor units within a human muscle. J. Neurol. Neurosurg. Psychiatry 1971; 34: 121-31.
- Kadhiresan VA, Hassett CA, Faulkner JA. Properties of single motor units in medial gastrocnemius muscles of adult and old rats. J. Physiol. (Lond) 1996; 493(Pt 2): 543-52.
- Kanda K, Hashizume K. Changes in properties of the medial gastrocnemius motor units in aging rats. J. Neurophysiol. 1989; 61: 737-46.
- Sugiura M, Kanda K. Progress of age-related changes in properties of motor units in the gastrocnemius muscle of rats. J. Neurophysiol. 2004; 92: 1357-65.
- Doherty TJ, Brown WF. The estimated numbers and relative sizes of thenar motor units as selected by multiple point stimulation in young and older adults. Muscle Nerve 1993; 16: 355-66.
- Doherty TJ, Vandervoort AA, Taylor AW, Brown WF. Effects of motor unit losses on strength in older men and women. J. Appl. Physiol. 1993; 74: 868-74.
- Holloszy JO. Workshop on Sarcopenia: Muscle Atrophy in Old Age. J. Gerontol. 1995; 50A: 1-161.
- Ishihara A, Naitoh H, Araki H, Nishihira Y. Soma size and oxidative enzyme activity of motoneurones supplying the fast twitch and slow twitch muscles in the rat. Brain Res. 1988; 446: 195-8.
- Tomlinson BE, Irving D. The numbers of limb motor neurons in the human lumbosacral cord throughout life. J. Neurol. Sci. 1977; 34: 213-9.
- Kanda K, Hashizume K, Nomoto E, Asaki S. The effects of aging on physiological properties of fast and slow twitch motor units in the rat gastrocnemius. Neurosci. Res. 1986; 3: 242-6.
- Andersson AM, Olsen M, Zhernosekov D, Gaardsvoll H, Krog L, Linnemann D, Bock E. Age-related changes in expression of the neural cell adhesion molecule in skeletal muscle: a comparative study of newborn, adult and aged rats. Biochem. J. 1993; 290 ( Pt 3): 641-8.
- Brown MC, Holland RL, Hopkins WG. Motor nerve sprouting. Annu. Rev. Neurosci. 1981; 4: 17-42.
- Wilson GJ, Newton RU, Murphy AJ, Humphries BJ. The optimal training load for the development of dynamic athletic performance. Med. Sci. Sports Exerc. 1993; 25: 1279-86.
- Svantesson U, Grimby G, Thomee R. Potentiation of concentric plantar flexion torque following eccentric and isometric muscle actions. Acta Physiol. Scand. 1994; 152: 287-93.
- Verdery RB. Failure to thrive in old age: follow-up on a workshop. J. Gerontol. Med. Sci. 1997; 52A: M333-M336.
- Lopez ME, Zainal TA, Chung SS, Aiken JM, Weindruch R. Oxidative stress and the pathogenesis of sarcopenia. In Sen CK, Packer L and Hanninen O. (eds). Handbook of Oxidants and Antioxidants in Exercise. Elsevier, Oxford. 2000.
ПЕРЕВОД ОСУЩЕСТВЛЯЛИ МАГИСТРАНТЫ:
Аннотация – Шершнева А.А.
Типы мышечных сокращений – Прошина В.А.
Слабость и неспособность развиваться – Андрейчик Е.О., Вуколова Е.В.
Механизмы, ответственные за атрофию скелетных мышц – Мищенко А.Д., Макаров Я.Н.
Механизмы, ответственные за потерю мышечных волокон – Козлов А.В.
Привычный уровень и тип физической активности – Матвеева М.С., Пивоварова Ю.В.
Общая редакция текста – Самсонова А.В.