Влияние силовых тренировок и возраста на изменения в силе и размере мышц
На силу и гипертрофию мышц человека влияет ряд факторов. Для точной оценки силы необходимо учитывать архитектуру мышц, влияние ЦНС, а также композицию мышечных волокон. Однако, даже учет этих факторов не может объяснить непропорционально большое изменение в силе, по сравнению с изменениями в объеме мышц после силовой тренировки или по мере старения человека.

Degens, R.M. Erskine, C.I. Morse Disproportionate changes in skeletal muscle strength and size with resistance training and ageing // J. Musculoskelet Neuronal Interact 2009; 9(3):123-129
Х. Дегенс, Р. М. Eрскин, С. И. Mорс
ВЛИЯНИЕ СИЛОВЫХ ТРЕНИРОВОК И ВОЗРАСТА НА НЕПРОПОРЦИОНАЛЬНЫЕ ИЗМЕНЕНИЯ В СИЛЕ И РАЗМЕРЕ СКЕЛЕТНЫХ МЫШЦ
Abstract
Способность мышцы сокращаться и производить силу играет решающую роль в передвижении, осанке, равновесии и дыхании. Во время сокращения головки миозина, нити миозина приводят в движение филаменты актина через гидролиз АТФ, в результате чего мышцы укорачиваются и/или развивается сила. Максимальная скорость укорочения мышечного волокна в значительной степени определяется активностью АТФ-фазы миозина, в то время как максимальная сила прежде всего определяется площадью поперечного сечения мышцы. Поскольку большинство мышц перистые, а не параллельные и работают при разных плечах силы, для получения характеристик силы и скорости мышцы, следует принимать во внимание строение мышц и анатомию суставов и сухожилий. Кроме того, активация антагонистических мышц (агонистов и антагонистов) будет способствовать развитию момента силы, генерируемого во времени сокращения. Наконец, соотношение сухожилий может повлиять на скорость нарастания силы. Усилие, создаваемое таким образом, позволяет сокращению мышц соответствовать восходящей ветви кривой «длина-напряжение». Несмотря на то, что для исследований связи изменения сократительных свойств в мышцах, изменений в размере мышц и строения в процессе силовых тренировок и старения применяются: магниторезонансная томография и УЗИ, в сочетании с ЭМГ и/или электростимуляцией, еще предстоит объяснить непропорциональное изменение мышечной силы и ее объема.
Ключевые слова: строение мышц, силовые тренировки, старение, сухожилия, относительное напряжение.
Введение
Скелетные мышцы играют важную роль в передвижении, осанке, равновесии и дыхании. Способность мышц сокращаться и генерировать силу, имеет решающее значение для выполнения этих функций. Однако, этой способности недостаточно для повседневной жизни.
Действительно, сила и скорость сокращения должны строго контролироваться, чтобы производить точные, четкие движения с минимальной силой, такие как продевания нитки в иглу, а также те, которые требуют максимальной силы и/или скорости, такие как поднятие тяжестей и бега.
Но что же контролирует сокращение мышц? Что определяет силу и скорость сокращения и что определяет максимальную силу генерирования мощности и скорости сокращения мышцы?
Эти вопросы задавали в течение многих лет и до сих пор они являются предметом многочисленных исследований. Разные условия, которые ассоциируются с непропорциональными изменениями в мышечной силе и их объеме, такие как силовые тренировки и возраст (Таблица 1), могут помочь пролить свет на факторы, которые определяют сократительные свойства мышц.
Контроль мышечного сокращения
Ян Сваммердам (Jan Swammerdam, 1667) был первым, кто показал, что раздражение нерва приводит к мышечному сокращению. Позже, Луиджи Гальвани (Luidgi Galvani, 1751) показал, что электрический импульс, перемещаясь по нервам вызывает мышечные сокращения (1). Теперь мы знаем, что деполяризация сарколеммы и распространение потенциала действия вдоль сарколеммы в Т-трубочки приводит к освобождению «Ca2+» из саркоплазматического ретикулума. Этот «Ca2+» связывается с тропонином «C» в актиновых нитях, вызывая конформационное изменение тропомиозина, что воздействует на актиновый центр миозина на актиновой нити и заканчивается сокращением мышцы. Весь этот процесс называется электромеханическим сопряжением, так как возбуждение нерва связано сокращением мышцы.
Исходя из приведенного выше описания можно сделать вывод, что любая стимуляция двигательного нерва заканчивается сокращением мышцы. Это действительно было бы так, если бы не то, что двигательный нерв состоит из пучка аксонов, имеющих свое происхождение из множества двигательных нейронов спинного мозга. Каждый двигательный нейрон возбуждает некоторое число мышечных волокон и потенциал действия, возникающий от двигательного нейрона, приводит к сокращению всех мышечных волокон, которые он иннервирует. Таким образом они работают как единое целое, отсюда термин двигательная единица (ДЕ). Такой механизм обеспечивает возможность контролировать силу и скорость сокращения, когда во время низкой интенсивности сокращения задействуются малые двигательные единицы, состоящие из медленных окислительных мышечных волокон, а с увеличением интенсивности больше и больше задействуются большие двигательные единицы, состоящие из быстрых волокон. Кроме того, тонкая настройка возможна за счет различной частоты раздражения и тем самым регулирования силы отдельных ДЕ, когда увеличение частоты раздражения приводит к увеличению силы. Очевидно, что такое нервно-мышечное взаимодействие, обеспечивает необходимую силу и скорость сокращения мышцы с нужным количеством силы и скорости, чтобы соответствующим образом выполнить определенную задачу.
Таблица 1. Изменения в максимальном произвольном напряжении (МVС) и поперечном сечении мышц (ППС) после силовой тренировки. Во второй части таблицы диспропорциональные изменения в МVС и ППC с возрастом.
| МVС,% | ППС,% | Пол | Возраст | Тип Трен. | Продолжит. | Мышца | Лит. |
| 15 | 5 | М, Ж | 27 | Изометрическая | 6 нед. | Сгибатели локтевого сустава | 7 |
| 36 | 7 | М, Ж | 22 | Изотоническая | 8 нед | Сгибатели локтевого сустава | 11 |
| 16 | 10 | М | 29 | Изотоническая | 16 нед. | Разгибатели локтевого сустава | 25 |
| 35 | 5 | М, Ж | 28 | Изометричеcкая | 12 нед. | Разгибатели коленного сустава | 10 |
| 11 | 5 | М, Ж | 28 | Эксцентрическая | 12 нед. | Разгибатели коленного сустава | 10 |
| 15 | 5 | М, Ж | 28 | Концентрическая | 12 нед. | Разгибатели коленного сустава | 10 |
| 30 | 19 | М | 29 | Изотоническая | 6 мес. | Разгибатели коленного сустава | 12 |
| 33 | 5 | М | 20 | Изотоническая | 9 нед. | Разгибатели коленного сустава | 8 |
| 39 | 7 | М, Ж | 20 | Изотоническая | 5 нед. | Разгибатели коленного сустава | 31 |
| 16 | 10 | М | 27 | Изотоническая | 14 нед. | Разгибатели коленного сустава | 6 |
| 36 | 5 | М | 42 | Изотоническая | 6 мес. | Разгибатели коленного сустава | 14 |
| 36 | 2 | М | 72 | Изотоническая | 6 мес. | Разгибатели коленного сустава | 14 |
| 66 | 9 | Ж | 39 | Изотоническая | 6 мес. | Разгибатели коленного сустава | 14 |
| 57 | 6 | Ж | 67 | Изотоническая | 6 мес. | Разгибатели коленного сустава | 14 |
| -47 | -161 | М | 25 против 74 | Икроножная мышца | 53 | ||
| -43 | -302 | Ж | 23 против 70 | Разгибатели коленного сустава; Сгибатели коленного сустава | 51 | ||
| -44 | -24 | М | 28 против 68 | Разгибатели коленного сустава | 63 | ||
| -32 | -14 | Ж | 28 против 68 | Сгибатели локтевого сустава | 63 | ||
| 1 Физиологический поперечник; 2 Мышечный объем | |||||||
Мышечная архитектура
Угол, под которым мышечные волокна прикрепляются к апоневрозу, (угол перистости), влияет на силу, действующую на сухожилие. Эта сила может быть вычислена, как произведение развиваемого усилия всех волокон на косинус их угла перистости, при условии, что апоневроз расположен параллельно сухожилию (3). Угол перистости также влияет как на скорость сокращения мышцы, так и на силу мышцы (4). Уменьшение скорости сокращении перистых волокон мышцы, однако, может быть компенсировано большим охватом движений мышечных волокон (хотя бы в течение начальных стадий сокращения), что может стать причиной более быстрого движения сухожилия по сравнению со скоростью сокращения мышечных волокон (5). Тем не менее, чем больше угол перистости, тем меньше сила и скорость сокращения, что приводит к снижению результирующей относительной силы (то есть силы, приходящейся на единицу мышечной массы) (Рисунок 1). На данном этапе можно задаться вопросом, какое превосходство имеют перистые мышцы над мышцами с параллельным расположением мышечных волокон? Объяснение может быть получено при рассмотрении сократительного элемента перистых мышц. После того как Ян Сваммердам ((Jan Swammerdam, 1664) открыл, что объем мышцы не изменяется во время сокращения, Нильс Стенсен (Niels Stensen, 1667) был первым, кто обратил внимание на то, что во время сокращения увеличение поперечного сечения мышцы должно соответствовать имеющейся площади поверхности апоневроза (3). К тому же, скорость сокращения всей мышцы меньше, чем скорость сокращения мышечных волокон, в связи с этим появляется возможность более точно управлять движениями.
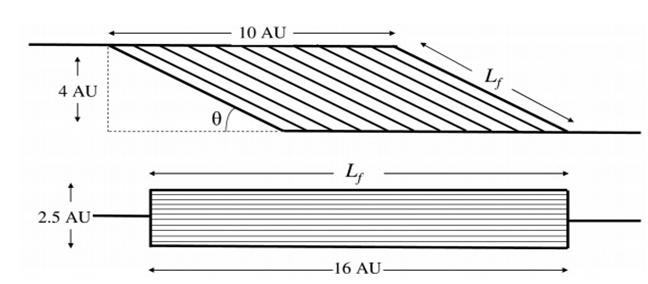
Сила, прикладываемая к сухожилию перистой и параллельной мышцы следующая:
- перистая: 5 у.е. (AU)*cos (30 град) = 4.33 у.е. (AU).
- параллельная: 2,5 у.е. (AU)*cos (0град) = 2,5 у.е. (AU).
Следовательно, перистая мышца производит в 1,73 (4,33 / 2,5) раза больше силы, по сравнению с параллельной мышцей.
Для подсчета скорости сокращения применяется следующее:
перистая: 8 у.е. (AU)*cos(30град) = 6.93 у.е. (AU);
параллельная: 16 у.е. (AU)*cos (0град) = 16у.е(AU).
Следовательно, перистая мышца сокращается в 0,43 (6,93/16) раза медленнее, чем параллельная мышца. Если перемножить эти два фактора (0,43 х 1,73) можно видеть, что перистые мышцы, производят только 74% от мощности параллельной мышцы того же объема.
Cиловые тренировки
Так как максимальная способность мышцы вырабатывать силу большей частью обусловлена количеством сакромеров, расположенных параллельно, можно предположить, что нарастание силы после силовой тренировки пропорционально увеличению объема мышцы. Из таблицы 1 следует, что кратковременные силовые тренировки вызывают большее увеличение силы мышцы, чем ожидалось от увеличения размеров данной мышцы у молодых (6-13) и пожилых людей (14-15). Ученые предположили, что такое непропорциональное увеличение мышечной силы имеет место в результате нервных влияний (11, 16), но данное явление наблюдается не всегда (10, 17, 18). Также возможно, что уменьшение взаимодействия мышц-антагонистов вызывает увеличение максимальных произвольных сокращений (момента силы) мышцы, которые возникают после силовой тренировки (14, 19, 20), хотя большинство ученых не наблюдают таких изменений во взаимодействии антагонистических мышц при тренировке (6, 21, 22). Кроме того, изменения в мышечной архитектуре (строении мышцы) могут стать причиной диспропорции между увеличивающимися силой и объемом мышцы.
Изменения в строении мышцы
Увеличение размера мышцы в ответ на силовую тренировку часто рассматривают как увеличение ее анатомического поперечника (ACSA), который оценивается посредством магнитно-резонансной или компьютерной томографией. Однако анатомический поперечник (ACSA), представленный как расстояние между соединительно-тканными образованиями на рисунке 1 не учитывает угол перистости (угол прикрепления мышечных волокон к апоневрозу в перистых мышцах). Более того, чем больше угол перистости, тем больше недооценивается значение физиологического поперечника (PCSA) в процессе измерения анатомического поперечника (ACSA) на основе измерения площади поперечного сечения мышцы (23, 24). Следовательно, благодаря сопутствующему увеличению угла перистости у молодых, старых и больных людей увеличение площади поперечного сечения мышечного волокна (fCSA) (6, 27-29), в результате тренировок не всегда вызывают увеличение анатомического поперечника мышцы (ACSA) (6). Положительным моментом увеличения угла перистости является то, к апоневрозу прикрепляется больше контрактильных волокон, что приводит к увеличению выработки силы мышцей (6, 30). Однако увеличение угла перистости может вызвать одновременное уменьшение силы мышечных волокон, связанных с апоневрозом пропорционально косинусу угла перистости (Рисунок 1). Несмотря на эти потери общий эффект влияния на силу, развиваемую сухожилием остается положительным, если угол перистости не превышает 45 градусов (23).
Некоторые исследователи наблюдали, что после силовой тренировки, длина мышечных волокон увеличивается как у юных (31), так и пожилых людей (22, 26), что позволяет увеличить максимальную скорость сокращения мышечного волокна. Так как максимальная мощность зависит от максимальной силы и максимальной скорости укорочения мышцы, увеличение длины волокна и физиологического поперечника (PCSA), должно привести к увеличению максимальной мощности, производимой мышцей. Тем не менее, увеличение угла перистости, вследствие силовой тренировки может не только уменьшить силу, развиваемую сухожилием, но и уменьшить максимальное укорочение мышцы в целом, тем самым ограничивая вызванное тренировкой увеличение силы и скорости сокращения мышцы (Рисунок 1). Сообщалось, что увеличение угла перистости в латеральной широкой мышце после силовой тренировки только на 2,7 градуса (6) приведет к потере силы, развиваемой сухожилием мышцы на 1%, это весьма незначительная потеря, учитывая, что максимальная сила была возрастает на 16% (6). Хотя большинство исследований действительно показывают увеличение выходной мощности мышц после силовой тренировки, как у юных (32, 33), так и у пожилых (34) людей, замедление сократительных свойств мышц, как следствие увеличения угла перистости, и сдвиг типа МВ с сторону более медленных (см. ниже) может объяснить, почему повышение силы, вызванное силовой тренировкой не обязательно сопровождается увеличением максимальной мощности (35).
Адаптации мышечных волокон
Изменения в мышечной силе и объеме после силовой тренировки, вероятно, сопровождаются изменениями в размере и фенотипе мышечных волокон. В самом деле, помимо увеличения площади поперечного сечения мышечного волокна (fCSA) (6, 16), силовая тренировка приводит к преобразованию мышечных волокон (27, 28), что отражается в сдвиге изоформ тяжелой цепи миозина (MHC) из типа IIx в IIa (28, 36, 37). Максимальная выходная мощность мышечного волокна IIa типа, тем не менее, меньше, чем у волокна IIx типа того же размера (38) и уменьшение процента мышечных волокон IIx та и увеличение процента мышечных волокон IIa типа может привести к уменьшению, а не увеличению мышечной силы в результате гипертрофии мышечных волокон. Другим фактором, который может объяснить непропорционально большое увеличение мышечной силы и объема в результате силовой тренировки является большая плотность упаковки миофибрилл в мышечных волокнах (39). Увеличение относительной силы (сила, деленная на площадь поперечного сечения мышечного волокна) (40, 41) поддерживает это соображение, но не соответствует исследованиям (42, 43).
В целом, ни изменения в нервном контроле, ни изменения мышечной архитектуры или качества волокон, по всей видимости, полностью не могут объяснить непропорциональное увеличение силы и мышечного объема. В связи с этим было высказано предположение, что это может объяснить поперечная передача силы от мышечных волокон к апоневрозам (44). Предполагается, что соединительно-тканные образования между промежуточными саркомерами и апоневрозом могут эффективно увеличить физиологический поперечник мышцы (PCSA), но уменьшить их функциональную длину. В результате мышца станет более сильной, но более медленной (44). Несмотря на то, боковая передача силы имеет место (45), это является предположением, что силовая тренировка вызывает изменения, которое могли бы объяснить непропорциональное увеличение мышечной силы и объема мышц.
Старение
В отличие от увеличения cилы в ответ на силовые тренировки, старение связано с прогрессивным ремоделированием нервно-мышечной системы, что имеет ряд последствий для силы мышц и в конечном счете для качества жизни. Саркопения (потеря массы, связанная с возрастом) хорошо документирована (46) и в значительной степени обусловлена снижением физической активности (47, 48) и потерей двигательных единиц, что является следствием уменьшения мотонейронов (49,50). Вклад саркопении в слабость пожилых людей может быть уменьшен снижением способности в полной мере рекрутировать агонисты во время силовых сокращений и снижением момента силы в результате повышенной совместной активации антагонистов (51, 52). Тем не менее, редко рассматривается влияние старения на изменение мышечной архитектуры и взаимосвязь изменений в силе и скорости сокращения относительно сухожилия.
Архитектура мышц и старение
Как уже обсуждалось выше, увеличение размеров мышц в ответ на силовые тренировки сопровождается изменением в мышечной архитектуре. Аналогичным образом, уменьшение объема мышц из-за старения также влияет на мышечную архитектуру. Уменьшение угла перистости и длины отдельных мышечных волокон (Lf) во время старения имеет ряд последствий для силы и скорости сокращения (53). Частично малый угол перистости у пожилых людей может компенсировать меньший физиологический поперечник мышцы (Рисунок 1). В действительности, однако, это теоретическое преимущество весьма незначительно. Действительно, уменьшение угла перистости на 2 градуса в икроножных мышцах пожилых людей дает только 1% преимущества в развитии максимального усилия (MVC) по сравнению с их молодыми коллегами (53). Однако на самом деле, (подобно об этом шла речь при описании силовых тренировок), нервная активация и архитектура мышц не полностью объясняют потерю силы во время старения (52, 53) и, скорее всего, отражают уменьшение силы отдельного мышечного волокна (54-57).
На первый взгляд, можно было бы ожидать, что снижение Lf в старости влияет на рабочую длину мышцы (Lm). Однако имеются исследования показывающие, что соотношение Lf к Lm остается постоянным во время старения для данной мышцы (53) и любое изменение физиологического поперечника (PCSA) (объем мышц делится на длину мышечного волокна Lf), поэтому пропорционально изменению объема мышц. Подразумевается, что, во-первых, мышечные волокна в пределах данной мышцы, при прочих равных условиях, всегда укорачиваются на тот же процент независимо от длины мышцы, что позволяет им работать в той же части зависимости «длина-сила»; во-вторых, это представляет собой естественную стратегию, позволяющую сохранить мощность и силу мышцы (так как последнее зависит от физиологического поперечника) за счет уменьшения скорости сокращения.
Свойства сухожилий
Старение связано с увеличением соответствия сухожильно-мышечного соединения и увеличением скорости укорочения мышечных волокон во время сокращения всей мышцы (сухожильно-мышечного комплекса) (58). Это может привести к большей степени укорочения волокон, чем было бы в случае, если сухожилия были жесткими. Если мышечные функции на восходящей части зависимости «длина-натяжение», такие как в икроножной и латеральной широкой мышцах (22), то большее укорочение мышечных волокон при сокращении мышцы может способствовать снижению связанного с возрастом мышечного напряжения (58). На 10% большее укорочение икроножной мышцы (59), может привести уменьшению на 20% относительного напряжения мышцы (53). Это явление проявляется во время подъема по лестнице, ходьбы и удержании равновесия, в частности, относительно пожилых людей, работающих гораздо ближе их максимальной мощности по сравнению с молодыми людьми (60-62). Было показано, что у пожилых людей, в результате повышения жесткости сухожилия надколенника, мышечные волокна латеральной широкой мышцы работают в более благоприятной части кривой «длина-напряжение» (22) и может, таким образом, компенсируют в какой-то степени связанную с возрастом потерю мощности мышц и медленность мышечно-сухожильного комплекса.
Заключение
Для того, чтобы получить точную оценку относительного напряжения и силы мышцы в естественных условиях, необходимо принимать во внимание мышечную архитектуру, нервное управление работой агонистов и мышц-антагонистов, а также композицию мышечных волокон. Однако, даже если мы примем во внимание эти факторы, непропорционально большее изменение в силе, чем в объеме мышц после силовой тренировки или по мере старения организма человека остается не полностью объясненным. Более детальные исследования необходимы для того, чтобы решить эту проблему путем тщательного рассмотрения широкого спектра факторов, определяющих мышечную силу и мощность, таких как нейронная архитектура управления и мышц. Считается, что архитектура мышц, оцениваемая с помощью ультразвука, дает двухмерное представление о мышце, в то время как мышечные волокна, особенно перистых мышц требуют трех измерений. Еще одним ограничением является то, что модели часто предполагают, что длина мышечных волокон и углы перистости одинаковы вдоль всей длины мышцы. Эти соображения в будущих экспериментах могут помочь объяснить непропорциональные изменения объема мышц и способность к генерации силы в результате силовых тренировок и старения. Также еще предстоит выяснить, является ли причиной этих диспропорций изменение качества мышечных волокон и / или увеличенные поперечные составляющие силы тяги мышцы.
Литература
1.Cobb M. Timeline: exorcizing the animal spirits: Jan Swammerdam on nerve function. Nat Rev Neurosci 2002; 3:395-400.
- Henneman E, Somjen G, Carpenter DO. Functional Significance of Cell Size in Spinal Motoneurons. J Neurophysiol 1965; 28:560-80.
- Huijing PA. Muscle, the motor of movement: properties in function, experiment and modelling. J Electromyogr Kinesiol 1998; 8:61-77.
- Narici M. Human skeletal muscle architecture studied in vivo by non-invasive imaging techniques: functional significance and applications. J Electromyogr Kinesiol 1999; 9:97-103.
- Gans C, Gaunt AS. Muscle architecture in relation to function. J Biomech 1991;24(Suppl.1):53-65.
- Aagaard P, Andersen JL, Dyhre-Poulsen P, Leffers AM, Wagner A, Magnusson SP, Halkjaer-Kristensen J, Simonsen EB. A mechanism for increased contractile strength of human pennate muscle in response to strength training: changes in muscle architecture. J Physiol 2001;534: 613-23.
- Davies J, Parker DF, Rutherford OM, Jones DA. Changes in strength and cross sectional area of the elbow flexors as a result of isometric strength training. Eur J Appl Physiol Occup Physiol 1988;57: 667-70.
- Erskine RM, Degens H, Jones DA. Factors contributing to an increase in quadriceps specific tension following resistance training in young men. Proc Physiol Soc 2008;11:C89.
- Ikai M, Fukunaga T. A study on training effect on strength per unit cross-sectional area of muscle by means of ultrasonic measurement. Int Z Angew Physiol 1970;28: 173-80.
- Jones DA, Rutherford OM. Human muscle strength training: the effects of three different regimens and the nature of the resultant changes. J Physiol 1987;391: 1-11.
- Moritani T, deVries HA. Neural factors versus hypertrophy in the time course of muscle strength gain. Am J Phys Med 1979;58: 115-30.
- Narici MV, Hoppeler H, Kayser B, Landoni L, Claassen H, Gavardi C, Conti M, Cerretelli P. Human quadriceps cross-sectional area, torque and neural activation during 6 months’ strength training. Acta Physiol Scand 1996;157: 175-86.
- Young A, Stokes M, Round JM, Edwards RH. The effect of high-resistance training on the strength and cross-sectional area of the human quadriceps. Eur J Clin Invest 1983; 13:411-7.
- Hakkinen K, Kallinen M, Izquierdo M, Jokelainen K, Lassila H, Malkia E, Kraemer WJ, Newton RU, Alen M. Changes in agonist-antagonist EMG, muscle CSA, and force during strength training in middle-aged and older people. J Appl Physiol 1998;84: 1341-9.
- Suetta C, Aagaard P, Rosted A, Jakobsen AK, Duus B, Kjaer M, Magnusson SP. Training-induced changes in muscle CSA, muscle strength, EMG, and rate of force development in elderly subjects after long-term unilateral disuse. J Appl Physiol 2004;97: 1954-61.
- Staron RS, Leonardi MJ, Karapondo DL, Malicky ES, Falkel JE, Hagerman FC, Hikida RS. Strength and skeletal muscle adaptations in heavy-resistance-trained women after detraining and retraining. J Appl Physiol 1991;70: 631-640.
- Komi PV, Buskirk ER. Effect of eccentric and concentric muscle conditioning on tension and electrical activity of human muscle. Ergonomics 1972;15: 417-34.
- Thorstensson A, Karlsson J, Viitasalo JH, Luhtanen P, Komi PV. Effect of strength training on EMG of human skeletal muscle. Acta Physiol Scand 1976;98: 232-6.
- Carolan B, Cafarelli E. Adaptations in coactivation after isometric resistance training. J Appl Physiol 1992;73: 911-7.
- Hakkinen K, Alen M, Kallinen M, Newton RU, Kraemer WJ. Neuromuscular adaptation during prolonged strength training, detraining and re-strengthtraining in middle-aged and elderly people. Eur J Appl Physiol 2000;83: 51-62.
- Aagaard P, Simonsen EB, Andersen JL, Magnusson SP, Halkjaer-Kristensen J, Dyhre-Poulsen P. Neural inhibition during maximal eccentric and concentric quadriceps contraction: effects of resistance training. J Appl Physiol 2000;89: 2249-57.
- Reeves ND, Narici MV, Maganaris CN. In vivo human muscle structure and function: adaptations to resistance training in old age. Exp Physiol 2004;89: 675-89.
- Alexander RM, Vernon A. The dimensions of knee and ankle muscles and the forces they exert. J Hum Mov Studies 1975;1: 115-23.
- Haxton HA. Absolute muscle force in the ankle flexors of man. J Physiol 1944;103: 267-73.
- Kawakami Y, Abe T, Kuno SY, Fukunaga T. Traininginduced changes in muscle architecture and specific tension. Eur J Appl Physiol Occup Physiol 1995;72: 37-43.
- Suetta C, Andersen JL, Dalgas U, Berget J, Koskinen S, Aagaard P, Magnusson SP, Kjaer M. Resistance training induces qualitative changes in muscle morphology, muscle architecture, and muscle function in elderly postoperative patients. J Appl Physiol 2008;105: 180-6.
- Campos GE, Luecke TJ, Wendeln HK, Toma K, Hagerman FC, Murray TF, Ragg KE, Ratamess NA, Kraemer WJ, Staron RS. Muscular adaptations in response to three different resistance-training regimens: specificity of repetition maximum training zones. Eur J Appl Physiol 2002;88: 50-60.
- Staron RS, Karapondo DL, Kraemer WJ, Fry AC, Gordon SE, Falkel JE, Hagerman FC, Hikida RS. Skeletal muscle adaptations during early phase of heavy-resistance training in men and women. J Appl Physiol 1994;76: 1247-55.
- Staron RS, Malicky ES, Leonardi MJ, Falkel JE, Hagerman FC, Dudley GA. Muscle hypertrophy and fast fiber type conversions in heavy resistance-trained women. Eur J Appl Physiol Occup Physiol 1990;60: 71-9.
- Gollnick PD, Timson BF, Moore RL, Riedy M. Muscular enlargement and number of fibers in skeletal muscles of rats. J Appl Physiol 1981;50: 936-43.
- Seynnes OR, de Boer M, Narici MV. Early skeletal muscle hypertrophy and architectural changes in response to high-intensity resistance training. J Appl Physiol 2007;102: 368-73.
- Aagaard P, Simonsen EB, Trolle M, Bangsbo J, Klausen K. Effects of different strength training regimes on moment and power generation during dynamic knee extensions. Eur J Appl Physiol Occup Physiol 1994;69: 382-6.
- Duchateau J, Hainaut K. Isometric or dynamic training: differential effects on mechanical properties of a human muscle. J Appl Physiol 1984;56: 296-301.
- Caserotti P, Aagaard P, Puggaard L. Changes in power and force generation during coupled eccentric-concentric versus concentric muscle contraction with training and aging. Eur J Appl Physiol 2008;103: 151-61.
- Rutherford OM, Greig CA, Sargeant AJ, Jones DA. Strength training and power output: transference effects in the human quadriceps muscle. J Sports Sci 1986;4: 101-7.
- Adams GR, Hather BM, Baldwin KM, Dudley GA. Skeletal muscle myosin heavy chain composition and resistance training. J Appl Physiol 1993;74: 911-5.
- Andersen JL, Aagaard P. Myosin heavy chain IIX overshoot in human skeletal muscle. Muscle Nerve 2000;23: 1095-104.
- Bottinelli R, Canepari M, Pellegrino MA, Reggiani C. Force-velocity properties of human skeletal muscle fibres: myosin heavy chain isoform and temperature dependence. J Physiol 1996;495: 573-86.
- Penman KA. Human striated muscle ultrastructural changes accompanying increased strength without hypertrophy. Res Q 1970;41: 418-24.
- Pansarasa O, Rinaldi C, Parente V, Miotti D, Capodaglio P, Bottinelli R. Resistance training of long duration modulates force and unloaded shortening velocity of single muscle fibres of young women. J Electromyogr Kinesiol 2008; [Epub ahead of print].
- Parente V, D’Antona G, Adami R, Miotti D, Capodaglio P, De Vito G, Bottinelli R. Long-term resistance training improves force and unloaded shortening velocity of single muscle fibres of elderly women. Eur J Appl Physiol 2008;104: 885-93.
- Trappe S, Williamson D, Godard M, Porter D, Rowden G, Costill D. Effect of resistance training on single muscle fiber contractile function in older men. J Appl Physiol 2000;89: 143-52.
- Widrick JJ, Stelzer JE, Shoepe TC, Garner DP. Functional properties of human muscle fibers after short-term resistance exercise training. Am J Physiol Regul Integr Comp Physiol 2002;283: R408-16.
- Jones DA, Rutherford OM, Parker DF. Physiological changes in skeletal muscle as a result of strength training. Q J Exp Physiol 1989;74: 233-56.
- Street SF. Lateral transmission of tension in frog myofibers: a myofibrillar network and transverse cytoskeletal connections are possible transmitters. J Cell Physiol 1983;114: 346-64.
- Lexell J, Taylor CC, Sjostrom M. What is the cause of the ageing atrophy? Total number, size and proportion of different fiber types studied in whole vastus lateralis muscle from 15- to 83-year-old men. J Neurol Sci 1988;84: 275-94.
- D’Antona G, Pellegrino MA, Carlizzi CN, Bottinelli R. Deterioration of contractile properties of muscle fibres in elderly subjects is modulated by the level of physical activity. Eur J Appl Physiol 2007;100: 603-11.
- Degens H, Alway SE. Control of muscle size during disuse, disease, and aging. Int J Sports Med 2006;27: 94-9.
- Lexell J. Evidence for nervous system degeneration with advancing age. J Nutr 1997;127: 1011 S-1013S.
- Tomlinson BE, Irving D. The numbers of limb motor neurons in the human lumbosacral cord throughout life. J Neurol Sci 1977;34: 213-9.
- 51.Macaluso A, Nimmo MA, Foster JE, Cockburn M, McMillan NC, De Vito G. Contractile muscle volume and agonist-antagonist coactivation account for differences in torque between young and older women. Muscle Nerve 2002;25: 858-63.
- Morse CI, Thom JM, Davis MG, Fox KR, Birch KM, Narici MV. Reduced plantarflexor specific torque in the elderly is associated with a lower activation capacity. Eur J Appl Physiol 2004;92: 219-26.
- Morse CI, Thom JM, Reeves ND, Birch KM, Narici MV. In vivo physiological cross-sectional area and specific force are reduced in the gastrocnemius of elderly men. J Appl Physiol 2005;99: 1050-5.
- D’Antona G, Pellegrino MA, Adami R, Rossi R, Carlizzi CN, Canepari M, Saltin B, Bottinelli R. The effect of ageing and immobilization on structure and function of human skeletal muscle fibres. J Physiol 2003;552: 499-511.
- Degens H. Age-related skeletal muscle dysfunction: causes and mechanisms. J Musculoskelet Neuronal Interact 2007;7: 246-52.
- Larsson L, Li X, Frontera WR. Effects of aging on shortening velocity and myosin isoform composition in single human skeletal muscle cells. Am J Physiol 1997;272:C638-49.
- Larsson L, Li X, Yu F, Degens H. Age-related changes in contractile properties and expression of myosin isoforms in single skeletal muscle cells. Muscle Nerve Suppl 1997;5: S74-8.
- Morse CI, Thom JM, Birch KM, Narici MV. Tendon elongation influences the amplitude of interpolated doublets in the assessment of activation in elderly men. J Appl Physiol 2005;98: 221-6.
- Maganaris CN. Force-length characteristics of the in vivo human gastrocnemius muscle. Clin Anat 2003;16: 215-23.
- Hortobagyi T, Mizelle C, Beam S, DeVita P. Old adults perform activities of daily living near their maximal capabilities. J Gerontol A Biol Sci Med Sci 2003;58:M453-60.
- Onambele GL, Narici MV, Maganaris CN. Calf muscletendon properties and postural balance in old age. J Appl Physiol 2006;100: 2048-56.
- Reeves ND, Spanjaard M, Mohagheghi AA, Baltzopoulos V, Maganaris CN. Older adults employ alternative strategies to operate within their maximum capabilities when ascending stairs. J Electromyogr Kinesiol 2009;19: 57-68.
- Klitgaard H, Mantoni M, Schiaffino S, Ausoni S, Gorza L, Laurent-Winter C, Schnohr P, Saltin B. Function, morphology and protein expression of ageing skeletal muscle: a cross-sectional study of elderly men with different training backgrounds. Acta Physiol Scand 1990;1 40:41-54.
Перевод осуществляли магистранты:
Ю.А. Кондакова – Abstract и Введение
П.А. Тютиков – табл.1, контроль мышечного сокращения
Е.Б. Котельникова – мышечная архитектура
А.А. Евсеева – силовые тренировки, изменения в мышечной архитектуре
К.А. Архипов – изменения в мышечной архитектуре, адаптация мышечных волокон
В.А. Краснов – старение, изменение мышечной архитектуры при старении
О.М. Савченко – свойства сухожилий, заключение
Общая редакция – А.В. Самсоновой
2009_Degens_Erskine_Morse_perevod.pdf