Физиологические изменения в скелетной мышце
В статье описываются физиологические изменения в скелетной мышце, связанные с тренировкой скоростно-силовых качеств. Выходная мощность мышцы определяется скоростью сокращения и силой мышц. Для увеличения мощности, необходимо увеличить либо скорость сокращения, либо мышечную силу, либо эти качества совместно.

D.A. Jones, 0.M. Rutherford and D.F. Parker Physiological changes in skeletal muscle as a result of strength training //Quarterly Journal of Experimental Physiology, 1989.– 74.– p.233-256.
Д.А. Джонс, О.М. Рутзерфорд, Д.Ф. Паркер
ФИЗИОЛОГИЧЕСКИЕ ИЗМЕНЕНИЯ В СКЕЛЕТНОЙ МЫШЦЕ КАК РЕЗУЛЬТАТ СИЛОВОЙ ТРЕНИРОВКИ (перевод статьи)
ВВЕДЕНИЕ
Составными частями успеха в любой физической активности, будь то спорт или бытовая деятельность, являются: мастерство, сила, скорость и выносливость. Объектом тренировки является соответствие сочетание этих составных частей с требованиями вида спорта и индивидуальными особенностями спортсмена. Настоящий обзор посвящен физиологическим изменениям, связанным с тренировкой скоростно-силовых качеств, таких как спринт, прыжки и метания, где главным объектом тренировки является увеличение выходной мощности. Выходная мощность мышцы определяется скоростью сокращения и силой мышц. Для увеличения выходной мощности, поэтому необходимо увеличить либо скорость сокращения, либо мышечную силу, либо эти качества совместно.
Изменения в скорости сокращения
Если повысить максимальную скорость ненагруженного сокращения мышц, то повысится и максимальная мощность (рис. 1). Скорость сокращения мышцы, зависит от композиции мышечных волокон, в то время как скорость сокращения отдельного волокна определяется ферментативными свойствами его актомиозиновых поперечных мостиков. Все мышечные волокна содержат генетическую информацию для производства быстрого или медленного миозина и других белков, и поэтому возможно, что медленные волокна могут быть преобразованы в быстрые в результате тренировки. Известно, что длительная, относительно низкая силовая активность, как, например, при длительной низкочастотной электрической стимуляции, приводит к изменениям, замедляющим скорость сокращения мышечных волокон (Salmons & Vrbova, 1969; Salmons & Henriksson, 1981; Rutherford & Jones, 1988).
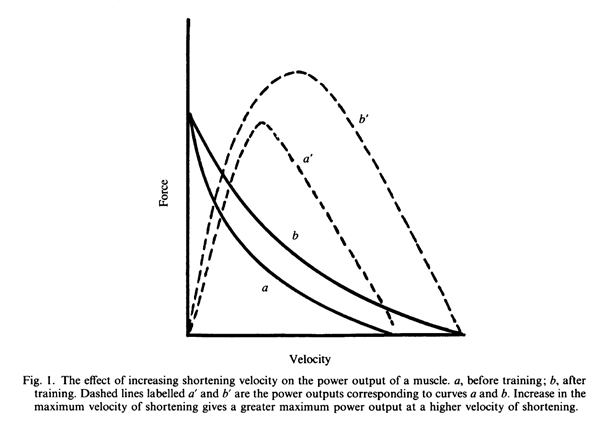
Опыт показывает, что обратное не верно. Длительная высокочастотная электрическая стимуляция мышц кошки вызвала такое же изменение их сократительных характеристик, какое наблюдалось при низкочастотной стимуляции (Eerbeek, Kernell и Verhey, 1984). Следовательно, кажется маловероятным, что тренировка будет улучшать мощность сокращения за счет увеличения скорости сокращения мышцы, вследствие изменения экспрессии генов. Существует, однако, другой способ увеличения скорости укорочения мышцы. Скорость сокращения мышцы зависит от скорости сокращения мышечных волокон, но также пропорциональна его длине (то есть количеству саркомеров, расположенных последовательно вдоль мышечного волокна). Если длина мышцы и соответственно число саркомеров увеличивается, например, в результате упражнений на растягивание, то максимальная скорость укорочения мышцы будет пропорционально увеличена. Эта возможность будет рассмотрена более подробно в разделе, посвященном специальной тренировке, воздействующей на длину и скорость сокращения мышцы.
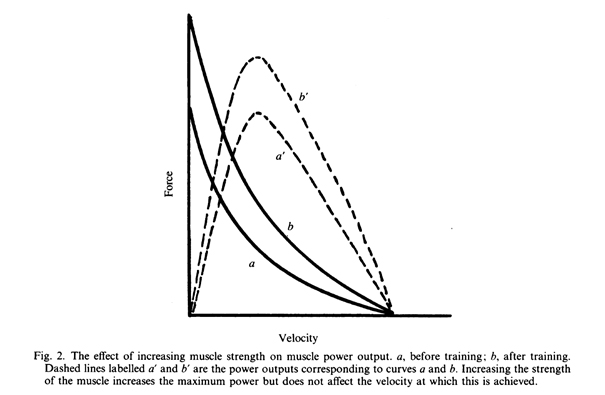
Изменения в силе мышцы
Если посредством тренировки невозможно увеличить скорость сокращения, тогда единственной альтернативой для тех, кто хочет увеличить выходную мощность является увеличение силы мышцы.
Увеличение максимальной изометрической силы, как ожидается, приведет к пропорциональному увеличению силы при всех скоростях сокращения и, следовательно, увеличению максимальной мощности (Рис. 2). В этом случае увеличение мощности не затрагивает скорость, при которой достигается максимальная мощность.
Силовая тренировка широко используется в качестве дополнения к обычной тренировке во многих видах спорта. Общая философия, поэтому следующая. Увеличивая размер и силу определенной группы мышц, увеличиваем выходную мощность в тех видах спорта, которые используют эти мышцы. Это, однако, довольно упрощенное представление.
История изучения силовой тренировки
В начале ХХ века Roux and Lange выдвинули теорию активной гипертрофии, предположив, что мышца выросла бы в размере и силе при необходимости, выполнить работу большей интенсивности, чем обычно (см. Hettinger, 1961). Это было ранней формой развитого позже принципа перегрузки deLorme (1946). В 1940-х и 1950-х годах капитан Tomas de Lorme, работающий с пациентами, после травм бедра и колена, развил идеи прогрессивного режима тренировки. Исследуемые тренировались с нагрузкой 60-90% от своего максимума, который оценивался еженедельно и, когда сила их мышц увеличивалась, масса груза увеличивалась. Ценность этой работы состояла в том, что была установлена важность работы высокой интенсивности, а не длительной работы низкой интенсивности. В 1956 году Hellebrandt and Houtz подтвердили это исследование, используя нормальных здоровых исследуемых, а не пациентов с существующей ранее атрофией мышц.
В 1950-х и 1960-х Hettinger and Muller выполнили ряд экспериментов, чтобы определить минимальный стимул, требуемый для увеличения силы мышц, также различные факторы, такие как возраст и пол, которые могли повлиять на способность к тренировке. Они пришли к заключению, что одно максимальное изометрическое сокращение продолжительностью 1-2 секунды в день было достаточно, чтобы произвести изменения в максимальной силе мышц. В ходе их исследований они заметили, что было широкое варьирование в восприимчивости различных исследуемых и мышц к тренировке. Это открытие продолжает интриговать исследователей и по сей день (Hettinger, 1961).
После этой ранней работы многие исследователи сравнивали эффекты изометрических, изокинетических и изотонических тренировок с разной комбинацией числа повторений, количество дней и недель тренировки, а также массу тренировочных грузов (см. обзоры Atha, 1981; McDonagh & Davies, 1984). Число и разнообразие этих исследований делают подробное сравнение почти невозможным, но основной вывод, который может быть сделан, состоит в том, что только десять повторений в день при нагрузках, больших, чем 60-70% от максимума при регулярном выполнении, произведут увеличение силы на 0,5-1% в день. Оригинальное заявление Hettinger and Mulller, что одно максимальное сокращение день достаточно для развития силы не было доказано другими исследователями (Bonde-Petersen, 1960), и даже сами авторы впоследствии изменили свои оригинальные заявления (см. Royce, 1964). Физиологическим основанием этих режимов является то, что при необходимости развития большого усилия активируются ДЕ, имеющие как низкие, так и высокие пороги активации (Milner Brown, Stein & Yemm, 1973) и поэтому они тренируются. Однако, сокращения мышц при более низкой интенсивности (приблизительно 30% от максимума), если проводятся более длительное время (60 с) чем обычно (2-5 с), как показывают эксперименты, также увеличивают уровень силы мышц (Davies &Young, 1983). В этом случае ДЕ, имеющие более высокий порог активации, вероятно, начинают работать в конце сокращения, когда другие уже устали.
Несмотря на интенсивные научные исследования и интерес к силовой подготовке, остаются неисследованные области, а также противоречия. Поэтому этот обзор будет иметь дело с тремя из этих областей.
Первая область – общее мнение, что тренировочный эффект очень специфичен относительно вида деятельности, скорости движения, а также длины активной мышцы.
Вторая область касается противоречия в том, что после тренировки прирост силы больше, чем ее размеров.
Заключительный аспект, который будет обсужден, касается природы стимула для гипертрофии мышц. Несмотря на очевидную важность этой темы, есть мало доказательств, на которых можно базировать рациональные тренировочные режимы для спортсменов и культуристов или для пациентов, подвергающихся восстановлению после ранения или болезни.
СПЕЦИФИКА ТРЕНИРОВОЧНЫХ ЭФФЕКТОВ
Очень часто в большинстве исследований указывается на то, что наибольшие изменения, сопровождающие силовые тренировки можно увидеть в самом упражнении, а не в параметрах какой-либо объективной оценки мышечной силы и ее размера (см., например, обзор Sale & MacDougall 1981).
Специфика задачи тренировки
Существует важное различие между силой, определяемой как способность выполнять задачу, которая может включать в себя скоординированные сокращения ряда мышечных групп и силой отдельных мышц. Тренировка часто проводится на силовых тренажерах, таких как (multi-gym). Опыт показывает, что способность поднимать тяжести дает большую прибавку, чем рост собственно силы мышечных групп тренирующихся (Hellebrandt и Houtz, 1956; Liithi, Howald, Claassen, Rosler, Vock и Hoppeler, 1986; Резерфорд и Джонс, 1986).
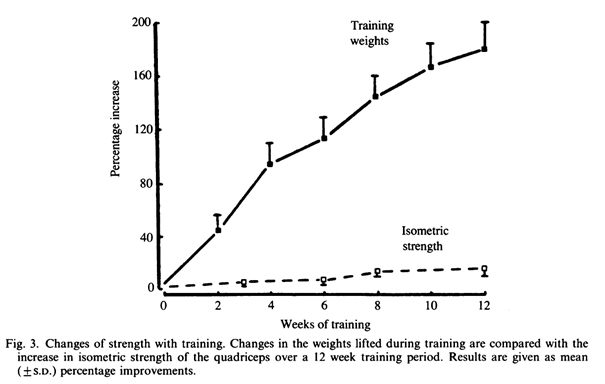
Rasch & Morehouse (1957) одними из первых сообщили о том, что эффект тренировки зависит от специфики движений. Субъекты тренировали силу сгибателей предплечья в положении стоя, а затем сила была измерена в положении стоя и лежа на спине. Увеличение мышечной силы в положении стоя было значительно больше, чем в незнакомом положении. Еще одним свидетельством специфики модели движения являются исследования изменения выходной мощности, которые сопровождали силовые тренировки. В одном исследовании испытуемые в течение 12 недель тренировались, в разгибании голени, поднимая почти субмаксимальные веса (Rutherford, Greig, Sargeant & Jones, 1986a). После трех месяцев тренировок, увеличение тренировочной нагрузки составляло порядка 200%, однако сопровождалось только 15% увеличением изометрической силы четырехглавой мышцы бедра (рис. 3). Изокинетическое оценивание выходной мощности, при котором используется модифицированный велоэргометр (Sargeant, Hoinville & Young, 1981), не показало никаких изменений в течение 12 недель. Несмотря на улучшение результатов тестирования, проводимых в знакомых тренировочных условиях, не было никаких существенных изменений в мощности, измеряемой в незнакомых условиях, хотя четырехглавая мышца является основной при выполнении движения на изокинетическом велоэргометре. Такое отсутствие перекрестного эффекта в тренировке предполагает, что значительное увеличение тренировочных весов может быть связано с приобретением навыка в задаче подъема тяжестей, которая имела минимальное значение при различных задачах педалирования на велосипеде.
Специфика длины мышцы
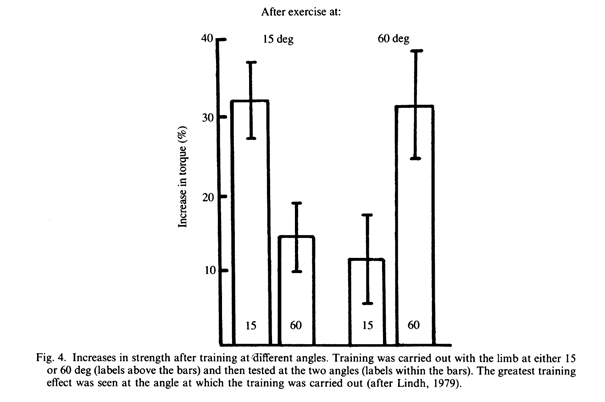
В ряде работ исследовались вопросы наращивания силы в результате тренировки при определенной длине мышц. Большинство исследователей обнаружили, что увеличение силы является наибольшим при той длине мышцы, при которой выполнялась тренировка. В одном из ранних исследований угла (Jablecki, 1975) и увеличенной радиологической плотности после шести недель тренировок людей, было обнаружено, что увеличение силы довольно специфично в зависимости от положения, в котором упражнялась конечность (Gardner, 1962). В более обширном исследовании Lindch (1979) тренировал испытуемых под углом в 15 или 60 град. в коленном суставе и проверял изометрическую силу мышц в обоих положениях. Было зафиксировано специфическое увеличение изометрической силы на 30% при положениях конечности, соответствующей углам тренировки. При положении конечности, соответствующей другим углам увеличение силы составляло только 12% (Рис. 4).
Концепция специфичности углов недавно была проверена для группы сгибателей локтевого сустава. В комплексном исследовании Thepaut-Mathieu, Hoecke&Maton (1988) проверялись изменения вследствие тренировки, под углом 120, 80 или 25 град в локтевом суставе. Наибольшая специфичность была после тренировки при укороченном состоянии мышцы (максимальном сгибании в локтевом суставе) (120 град.), наименьшая специфичность – в группе, которая тренировалась в удлиненном состоянии мышцы (разгибании в локтевом суставе). По результатам этого исследования можно предположить, что степень специфичности зависит от длины мышцы, при которой выполнялась тренировка: чем меньше длина, тем больше специфичность.
В противоположность указанным выше сообщениям, несколько авторов не нашли доказательства специфичности. Rasch, Pierson & Logan (1961) тренировали сгибатели локтевого сустава в изометрическом режиме под углом 90 град и обнаружили значительное и равное увеличение силы при углах в суставе в диапазоне от 45 до 135 град. Более позднее исследование Rasch & Pierson (1964) подтвердило более ранние наблюдения.
Специфичность, в любом исследовании, как правило, объясняется «нервной адаптацией» и возможности этого обсуждаются ниже. Однако, альтернативные объяснения, основанные на свойствах мышц, часто упускаются из виду, хотя могут играть важную роль. Первая возможность возникает из того факта, что группы мышц, такие как сгибатели предплечья или разгибатели голени состоят из нескольких отдельных мышц. Соотношения угла и напряжения у таких групп мышц определяются индивидуальными соотношениями длины и напряжения у мышц-компонентов. Различные мышцы могут иметь свой оптимум напряжения при различных углах между звеньями, тем самым расширяя полезный рабочий диапазон группы мышц (рис.5А). Если в результате тренировки была отмечена преимущественная гипертрофия одной из мышц, это может повлиять на соотношение угла и напряжения для всей группы мышц (рис. 5В). Таким образом, реакция на тренировку максимально проявится под углом, соответствующим оптимальной длине отдельной мышцы.
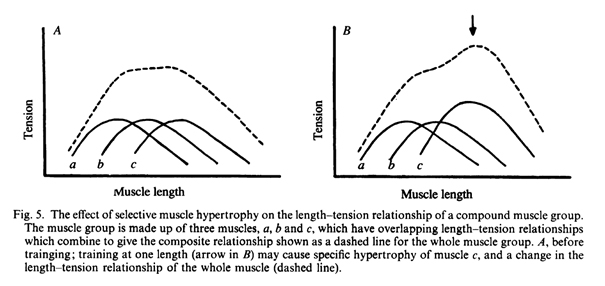
Вторая возможность возникает в результате изменения длины мышечных волокон. Если тренировка выполнялась при очень растянутой или очень укороченной мышце, когда силы, развиваемые мышечными волокнами намного ниже максимума, тогда может произойти удлинение либо укорочение мышцы путем добавления либо потери саркомеров в окончаниях мышечных волокон (Goldspink, Tabray, Tabray, Tardieu & Tardieu, 1974; Williams & Goldspink, 1978). Такое изменение в длине может сдвинуть соотношение длины и напряжения, так что максимальное перекрытие волокон с саркомерами, около 25 мкм, достигается, когда длина мышцы соответствует длине мышцы при тренировке (Рис. 6).
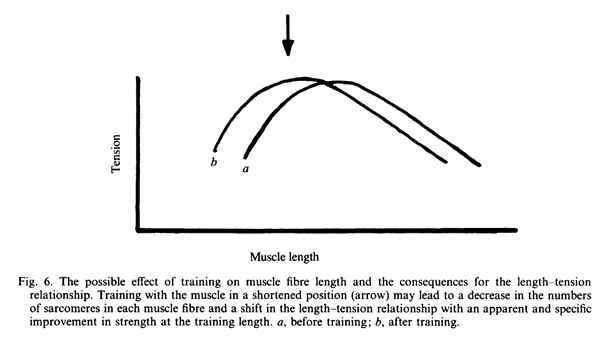
Имеются два способа проверить эти идеи. Один состоит в том, чтобы измерить изменения в размере отдельных мышц внутри группы мышц, такой как четырехглавая мышца, чтобы увидеть, имеются ли какие-нибудь свидетельства преимущественной гипертрофии одной или более мышц во время тренировки. Использование ЯМР [ядерно-магнитного резонанса] в настоящее время делает это возможным, и предварительные результаты говорят о том, что у четырехглавой мышцы может наблюдаться различная степень гипертрофии в ее головках (Cerretelli, Minetti, Narici, Roi & Scalmani, 1989). Второй подход может состоять в измерении соотношения длины и напряжения у одной из мышц-синергистов. Если изменится количество саркомеров – это проявится в изменении соотношения «длина-напряжение» мышцы. Тогда как если это было просто преимущественной гипертрофией, там не должно быть такого изменения. Обе группы мышц, четырехглавая и сгибатели предплечья, включают в себя мышцы, которые обслуживают два сустава (rectus femoris и biceps brachii caput medialis), которые позволяют соотношениям длины и напряжения в этих мышцах быть определенными независимо от других мышц в той же группе (Herzog & ter Keurs, 1988a, b).
Специфичность скорости
Проблема специфичности скорости в силовой тренировке в последнее время привлекла большое внимание, особенно с появлением в широком доступе изокинетических тренажеров фирмы Cybex. Скелетные мышцы человека отражают тесную взаимосвязь между силой и скоростью, как было описано во множестве исследований (Wilkie, 1950; Close, 1972; Thomas, White, Sagar & Davies, 1987), в связи с чем можно сделать вывод, что рост изометрической силы мышц вызовет рост силы в разных диапазонах скорости. Тем не менее, так считалось не всегда. Одно из самых ранних исследований изокинетического тренинга было сделано Moffroid и Whipple (1970), которые тренировали испытуемых на высоких и низких скоростях. Группа, тренировавшаяся на низких скоростях, по результатам тестирования показала значительный прирост результатов только на низких скоростях, в то время как группа, тренировавшаяся на высоких скоростях, показала однородный рост результатов на всех скоростях, и это было подтверждено в работах Coyle, Feiring, Rotkis, Cote, Roby, Lee и Wilmore (1981).
Caiozzo, Perrine и Edgerton (1981) и Kaneshi и Miyashita (1983) доказали наибольший прирост силы на скорости, приближенной к скорости во время тренировок (рис. 7A и B). Lesmes, Costill, Coyle и Fink (1978) отметили значительный рост силы на скорости, используемой во время тренировок, и также более высокой. Данные, подтверждающие концепцию специфичности скорости в тренировке, не являются до конца исчерпывающими. Следует с осторожностью рассматривать данные, полученные в результате коммерческих исследований изокинетических тестовых тренажеров, так как выводы о взаимосвязи между силой и скоростью очень условны и имеют мало общего с результатами, полученными при изолированном тестировании мышц (Close, 1972) и в условиях in situ (Wilkie, 1950). Казалось бы, тренировка отражается в скачках кривых сила-скорость (рис. 7), что, опять же, находится в противоречии с традиционными обоснованиями взаимосвязей. Причиной этих расхождений, скорее всего, являются недостатки частотных характеристик систем записи крутящего момента, которые могут внести значительные неточности в полученные результаты (см. Thomas и др., 1987 для обсуждения).
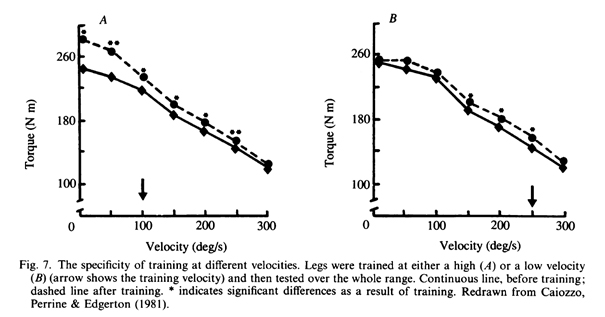
роме того, в то время как некоторые исследования фиксируют крутящий момент при конкретных углах, другие фиксируют пиковый крутящий момент при затухающем сигнале, что может скрывать истинные пиковые значения. Важно рассмотреть механизмы, которые могут привести к эффекту специфичности скорости, оставляя в стороне возможные нервные адаптации.
Изменение в силе мышечного волокна изменит максимальную скорость сокращения мышцы. Нет причины думать, что это произойдет в результате тренировки на высокой или низкой скоростях, но эта вероятность должна быть проверена путем изучения взаимосвязи между длиной и напряжением мышцы, отражающей эффект специфичности скорости. При быстром сокращении медленные мышечные волокна могут быть пассивны, и потому они либо будут производить очень низкую скорость, либо не будут производить ее вообще. Если скорость сама по себе является стимулом для гипертрофии, то высокоскоростные сокращения могут обеспечить стимул для роста «быстрых» двигательных единиц, что, возможно, объясняет специфику высокоскоростной тренировки. Тем не менее, нельзя провести ту же аналогию с «медленными» двигательными единицами: нельзя утверждать, что все они будут задействованы и получат стимул к гипертрофии.
Нервная адаптация
Возможно, что нетренированные испытуемые не в состоянии простым однократным усилием полностью активировать те или иные группы мышц с определенной (высокой) скоростью или с фиксацией конечности в определенном положении. Нет сомнения, что во время быстрых, резких движений испытуемый не может быть уверен, что усилие действительно максимально, и сокращения мышц, конечно, ощущаются весьма необычными, если пытаться фиксировать руку или ногу в полностью согнутом или разогнутом положении. Несомненно, тренировка может помочь испытуемому полностью активировать ту или иную мышцу в полном спектре движений и скоростей, но существует мало прямых доказательств, что это — объяснение специфики изменения длины и скорости сокращения, и было бы правильнее считать верными объяснения, приведенные выше, основанные на функционировании мышц
Специфика задачи здесь может состоять в улучшении координации различных групп мышц, которые вовлечены в такие действия, как тренировка езде на велосипеде или написание текста недоминирующей рукой. Нервные импульсы, вовлеченные в такие простые, на первый взгляд, действия, на самом деле весьма сложны и могут существовать на нескольких уровнях центральной нервной системы и включать различные сенсорные импульсы от кожи, суставов, глаз и вестибулярного аппарата. Также многие задачи тренировочного процесса, как может показаться, требуют минимального умения, но все это весьма относительно. Наблюдение за испытуемыми, которые с легкостью могут активизировать почти любую мышцу, показывает, что им также требуется сокращение многих различных мышц для фиксации в определенном положении конкретной конечности или части тела. Так сокращение мышц брюшной полости и мышц стенки грудной клетки считается универсальным и необходимым для осуществления большинства максимальных усилий. Это является частью приобретенного умения, которое должно перейти в навык.
ИЗМЕНЕНИЯ В МЫШЕЧНОЙ СИЛЕ И РАЗМЕРАХ ВО ВРЕМЯ ТРЕНИРОВКИ
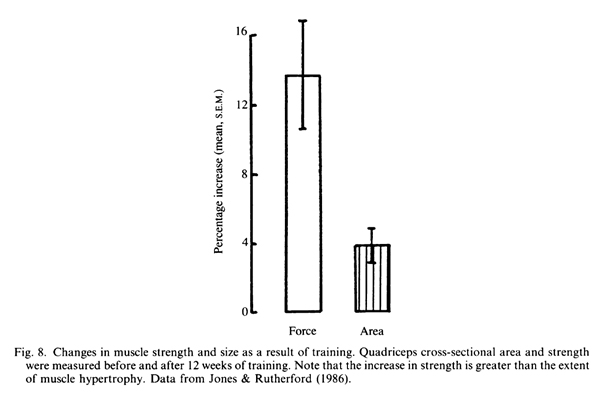
Краткосрочные наблюдения за тренировочным процессом продемонстрировали увеличения силы, которые оказались больше, чем увеличения размера мышц (Ikai & Fukunaga, 1970; Moritani & deVries, 1979; Young, Stokes, Round & Edwards, 1983; Jones & Rutherford, 1987; Рис. 8).
Нервные механизмы
Утверждалось, что до тренировки, мышцы не могут быть максимально активизированы при произвольном напряжении. В первые шесть — восемь недель тренировок до изменений в размерах мышц становится очевидным предположение, что активация, и, следовательно, сила увеличивается в результате изменения нервного контроля. Данные, подтверждающие эту точку зрения, следуют из записи поверхностной ЭМГ, где было продемонстрировано увеличение примерно на 10% максимума интегрированной ЭМГ в результате тренировок (Moritani & deVries, 1979; Komi, 1986).
Существуют однако, и противоречивые факты, свидетельствующие об отсутствии изменений, либо уменьшении максимальной активации с тренировкой (Komi & Buskirk, 1972; Thorstensson, Karlsson, Viitasalo, Luhtanen & Komi,1976b).
Существуют два основных способа, посредством которых нервная адаптация может привести к увеличению максимальной произвольной изометрической силы.
(1) Большие быстрые двигательные единицы рекрутируются только при необходимости развить максимальную силу, и возможно, что во время максимальных произвольных сокращений есть некоторые двигательные единицы, которые никогда не рекрутируются у нетренированных лиц. Тренировка, поэтому, рассматривается как способ облегчения рекрутирования этих больших и быстрых двигательных единиц.
(2) Характер электрической стимуляции ДЕ может измениться. Электрическая стимуляция мышц человека, через двигательный нерв или периферические ветви, требует частоты около 50-100 Гц для создания максимальной тетанической силы, но измерения частот естественных сокращений ДЕ, предполагает, что частота 20 Гц близка к физиологическому максимуму. Это различие может быть объяснено по результатам Rack & Westbury (1969) и Lind и Petrofsky (1978), которые показали, что асинхронная стимуляция при относительно низкой частоте частей отдельных нервов дает больше напряжение, чем синхронные стимуляции на той же частоте.
Объяснение этого явления, вероятно, касается внутренней работы, выполняемой мышцами против последовательного упругого компонента. При синхронной стимуляции на низких частотах все ДЕ будут активированы одновременно, и по мере развития напряжения они будут растягивать последовательный упругий компонент. При этом мышцы будут сокращаться, и, следовательно, будут развивать силу меньше, чем максимальное изометрическое усилие. При асинхронной стимуляции последовательный упругий компонент будет растягиваться первой частью мышечных волокон, затем он будет растягиваться второй и последующими частями мышечных волокон и генерировать полную силу. Komi (1986), однако, представил доказательства того, что у тренированных мышц существует увеличение силы при синхронизации. Пока не ясно, каким образом это изменение привело бы к увеличению изометрической силы, но это может быть функционально важно, когда необходимо быстро развивать максимальную силу, например, во время прыжков или метаний.
На основе записи ЭМГ можно зарегистрировать две детали. Ранее не активные рекрутированные ДЕ повышают частоту ЭМГ или увеличивают синхронизацию, что в свою очередь увеличивает максимум интегрированной ЭМГ. Асинхронная активность ДЕ приводит к снижению интегрированной ЭМГ, что является результатом индивидуальных потенциалов этих групп мышц, которые перекрывают друг друга во время записи на поверхностных электродах.
Есть ряд технических проблем связанных с записью ЭМГ активности во время длительных тренировок. ЭМГ сигнал, получаемый с поверхности электродов, во многом зависит от их положения на коже, а так же от сопротивления мышечных тканей и кожного покрова. Ученые стараются поместить электроды в такие места, где сопротивление кожи минимально. Однако данный процесс связан с серьезными трудностями ввиду того, что невозможно предугадать содержание межтканевого и внутримышечного жира, содержание которого может варьироваться в процессе длительного исследования.
Перекрестный тренировочный эффект наблюдается в ряде исследований, в которых нагрузка на одну конечность привела к увеличению силы мышц нетренируемой конечности. Сторонники «нейронной гипотезы» положили в основу утверждение, что этот эффект является результатом адаптации, влияющим на обе конечности.
Увеличение силы с помощью перекрестного тренировочного эффекта в пределах от 10 до 30% было продемонстрировано целым рядом ученых (Darcus & Salter, 1955; Komi, Viitasalo, Rauramara & Vihko, 1978; Moritani & deVries, 1979), тогда как другие группы исследователей не нашли никаких доказательств такого эффекта (Kruse & Mathews, 1955; Young et al. 1983; Jones & Rutherford, 1987; Davies, Parker, Rutherford & Jones, 1988). Young et al. (1983) предположили, что эти противоречивые результаты можно объяснить активностью, нетренируемой конечности во время тренировок. Если бы эта теория была верна, то мышечные волокна сокращались так же и в нетренируемой конечности, получая сигнал от тренируемой, что не возможно.
Согласно другому подходу к этому вопросу, произвольное сокращение полностью активных мышц можно увеличить до максимума, чтобы понять, сколько дополнительной силы можно получить (Merton, 1954; Belanger & McComas, 1981; Bellemare & Bigland-Ritchie, 1984; Gandevia & McKenzie, 1985; Rutherford, Jones & Newham, 1986b; Рис.9 9). Общим же заключением стало то, что человек может активировать различные группы мышц без каких-либо тренировок. В нашем эксперименте мы проверяли людей до, во время и после тренировок, при этом был обнаружен лишь один человек, у которого была выявлена двигательная аномалия (Jones & Rutherford, 1987). Сложно представить, в какой степени могут быть развиты мышцы для выявления максимального показателя силы.
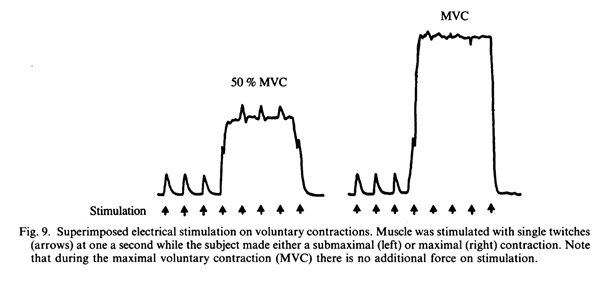
В связи с тем, что измерения тетанической силы вызывают сложности и неудобства, связанные с необходимостью сверхмаксимальной тетанической стимуляции двигательного нерва человека, исследований в этом направлении было проведено очень мало. Результаты нескольких исследований, которые были сделаны, противоречивы. McDonagh, Hayward & Davies (1983), обнаружили, что пять недель тренировки с максимальным изометрическим сокращением двуглавой мышцы плеча привели к увеличению произвольной силы на 20% без изменения тетанической силы при стимуляции с частотой в 40 Гц. В той же группе найдено, что восемь недель изометрической тренировки первой дорсальной межкостной мышцы (FDI) вызвало увеличение на 33% произвольной силы, но тетаническая сила не изменилась (Dooley, McDonagh & White,1983). В связи с этим авторы пришли к выводу, что увеличение нервного управления было ответственно за увеличение произвольной силы мышц. Однако в первом из этих исследований двуглавой мышцы начальная тетаническая сила была значительно ниже, чем произвольная сила. На основе этого было выдвинуто два предположения: 1) вся мышца не стимулировалась или 2) другие группы мышц способствовали произвольному сокращению. Во втором исследовании по FDI значения тетанической силы мышцы не сообщались. В ходе аналогичного исследования Duchateau & Hainaut (1984) обнаружилось, что три месяца изометрической или динамической тренировки мышцы, отводящей большой палец, привело к увеличению тетанической силы на 20% и 11% соответственно. Полученные значения очень похожи на те, которые нашли другие исследователи для увеличения произвольной силы.
Периферические механизмы
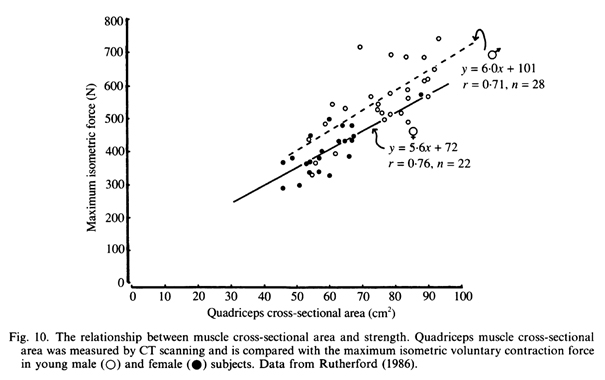
Альтернативные объяснения несоответствия между изменениями в мышечной силе и размером мышцы после тренировки основываются на периферических изменениях в составе мышцы, приводящих к более высокому специфическому напряжению. Ниже рассматривается ряд возможных механизмов, и становится очевидным, что в этой области имеются две основные проблемы: 1) методологические трудности в изучении состава мышцы в организме человека и 2) отсутствие понимания факторов, которые определяют силу мышц в неподготовленном состоянии. Только около 50% вариации в силе четырехглавой мышцы между людьми могут быть объяснены различием в площади поперечного сечения мышц (Chapman, Grindrod & Jones, 1984; Rutherford, 1986; рис.10). Другие факторы, которые могут повлиять на силу, включают в себя систему рычагов, через которую это сила измерена (McCullough, Maughan, Watson & Weir, 1984), тип МВ (Young,1984; Grindrod, Round & Rutherford, 1987), структуру волокна (Alexander & Vernon, 1975) и упаковку сократительного материала (Penman, 1970). Вполне возможно, что последние три могут быть изменены в период силовых тренировок.
Композиция мышечных волокон
Существуют доказательства, что при движениях человека и животных, что МВ II типа проявляют большую силу, чем первого. Исследования человеческой мышцы показали, что есть различия и в волокнах второго типа (Komi, Rusko, Vas & Vihko, 1977; Tesch & Karlsson,1978; Young,1984; Grindrod et al.,1987), но есть также противоречивые сообщения, показывающие, что тип мышечных волокон не влияет на силу мышц (Thorstensson et al.1976b; Clarkson, Kroll & McBride,1980; Maughan & Nimmo,1984). Результаты исследования на животных также являются противоречивыми, однако некоторые группы исследователей находят, что МВ II типа проявляют большую силу (Barany & Close,1971;Burke & Tsairus,1973; Kean,Lewis & McGarrick,1974; Edjtehardi & Lewis, 1979). Если МВ II типа имеют большую силу чем МВ I типа первого типа, то гипертрофия МВ II типа после тренировки приведет к большему увеличению мышечной силы, по сравнению с площадью поперечного сечения в смешанной мышце. Не смотря на то, что у элитных атлетов было найдено больше волокон второго типа (McDougall, Sale, Elder & Sutton,1982; Tesch & Karlsson,1982), доказательства этого в краткосрочных исследованиях тренировки противоречивые (Thorstensson et al.1976b; MacDougall, Elder, Sale, Moroz & Sutton,1980; Young et al.1983; Rutherford,1986). Различия в найденной гипертрофии были слишком малы, чтобы учитывать изменения в силе и в размере всей мышцы. Не существует каких-либо доказательств о преобразовании типов волокон после силовой тренировки (см. обзор Edstrom & Grimby,1986). Некоторая осторожность необходима в выводах, полученных на основе биопсии мышц человека, поскольку есть большие индивидуальные различия в гипертрофии мышечных волокон, хотя режим тренировки был одним и тем же. В нашем опыте изменения в площади поперечного сечения мышечных волокон может колебаться от -13 до +47%. Этот вывод подтверждается другими исследованиями (Yong et al., 1983; MacDougall,1986).
Угол прикрепления мышечных волокон
Существует большая вариация мышц по расположению мышечных волокон между сухожилиями. Самая простая архитектура – это архитектура, в которой мышечные волокна расположены параллельно сухожилиям (мышцы с параллельным расположением пучков мышечных волокон). Многие мышцы, включая четыре части четырехглавой мышцы бедра, являются волокнами перистой мышцы, и соединяются с сухожилием под углом к линии действия мышцы (угол перистости). В этой ситуации площадь поперечного сечения мышцы, измеренная под прямым углом к мышце (анатомический поперечник), недооценивает истинное поперечное сечение самих волокон (физиологический поперечник) (Edgerton, Roy & Apor, 1986). Изменение угла перистости может изменить силу, измеренную между концами мышцы. При той же самой длине и анатомическом поперечнике мышцы увеличенный угол перистости может привести к большему количеству контрактильного материала, прикрепленного к более крупной области сухожилия. Отношения между силой сухожилия, углом перистости и суммой контрактильного материала, сложны. Alexander & Vernon (1975) вычислили, что эта сила пропорциональна синусу двойного угла перистости. Эта модель предсказывает, что сила мышцы будет увеличиваться пока угол перистости не достигнет 45 град. У человека угол перистости четырехглавой мышцы бедра составляет приблизительно 13-18 град. Если бы в результате тренировки произошло одновременное увеличение и угла перистости и площади поперечного сечения мышечных волокон, то полное изменение площади поперечного сечения мышц было бы меньшим, чем площади волокна. Сейчас невозможно измерить точно угол перистости в естественных условиях, но косвенный способ исследовать эту проблему состоял бы в том, чтобы изучить изменения, происходящие в мышце с параллельным ходом пучков мышечных волокон после тренировки. В мышце с параллельным ходом пучков мышечных волокон физиологический поперечник может быть оценен на основе анатомического поперечника. Основные сгибатели предплечья являются мышцами с параллельным ходом пучков мышечных волокон, поэтому изучение результатов тренировки этих мышц должны дать качественно и количественно результаты, подобные тем, в которых предполагалось, что изменение в архитектуре волокна не является главной причиной увеличения мощности мышцы после тренировки.
Упаковка сократительного материала
Увеличение силы на единицу площади может быть объяснено повышенным упаковки сократительного материала. Это возможно из-за более плотной упаковкой миофиламентов или миофибрилл или потери жира и соединительной ткани между волокнами. В мышцах животных было обнаружено увеличение плотности миофиламентов (Helander, 1961) и миофибрилл (Gordon, Kowalski & Fritts, 1967) после периода силовой тренировки, хотя различия в плотности мышц как сообщается, были очень маленькими по сравнению с мышцами нетренированных животных (Mendez & Keys, 1960). Результаты исследований на людях противоречивы: MacDougall, Sale, Moroz, Elder, Sutton & Howald (1979) обнаружили, что силовая тренировка приводит к увеличению количества и размеров миофибрилл, но это сопровождалось значительной гипертрофией мышечного волокна, так что плотность миофибрилл осталась прежней. Наоборот, в исследовании с тремя испытуемыми, Penman (1970) обнаружил, что силовые тренировки привели к увеличению на 50% плотности нитей миозина без изменения обхватов конечностей. Было зафиксировано одновременное увеличение мышечной силы на 40%, которое он отнес к увеличению плотности упаковки миозина. В нескольких исследованиях на основе компьютерной томографии (КТ) было обнаружено увеличение радиационной плотности мышц после тренировки (Horber, Scheidegger, Grunig & Frey, 1985; Jones & Rutherford, 1987). Это может происходить по ряду причин: уменьшения содержания жира в мышцах, увеличения упаковки сократительных элементов или увеличения содержания соединительной ткани. Следствием первых двух вариантов будет увеличение силы на единицу площади. Косвенным свидетельством более плотной упаковке мышечных волокон является увеличение после тренировки площади поперечного сечения мышечных волокон по сравнению площадью поперечного сечения мышцы (McDonagh & Davies, 1984; Rutherford, 1986; Frontera, Meredith, O’Reilly, Knuttgen & Evans, 1988). В среднем увеличение площади поперечного сечения мышц составляет 5-10% (Dons, Bollerup, Bonde-Petersen & Hancke, 1979; Young et al. 1983; Luthi et al. 1986; Jones & Rutherford, 1987; Cerretelli et al. 1989) в то время как увеличение площади поперечного сечения мышечных волокон ближе к 15-20% (MacDougall, Sale, Moroz, Elder, Sutton & Howald, 1979; MacDougall et al. 1980; Rutherford, 1986; Frontera et al. 1988). Тем не менее, как обсуждалось выше, это несоответствие может объясняться перестройкой архитектуры волокна.
Прикрепление соединительных тканей
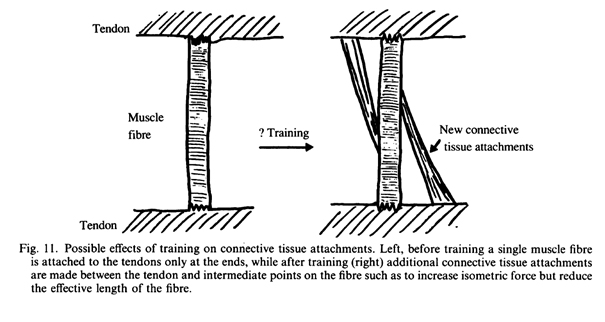
Принято считать, что напряжение в мышечных волокнах передается в продольном направлении через последовательный ряд саркомеров таким образом, что сила пропорциональна только площади поперечного сечения и не зависит от длины мышцы. Если, однако, между промежуточными саркомерами и соединительной тканью есть зоны прикрепления, это приведет к увеличению силы, создаваемой на единицу поперечного сечения мышцы (рис. 11). Мышечные волокна млекопитающих окутаны соединительной тканью, которая может играть определенную роль в передаче напряжения сухожилию (Street, 1983). Как известно, гипертрофия, вызванная тренировками, приводит к увеличению синтеза коллагена в мышцах животных (Schiaffino, Bornioli & Aliosi, 1972; Goldberg, Etlinger, Goldspink & Jablecki, 1975) и увеличенная радиологическая плотность, найденная у человека после тренировки (Horber et al. 1984; Jones & Rutherford, 1987) может быть результатом повышенного содержания соединительной ткани (Grindrod et al., 1982).
Последствия для выходной мощности
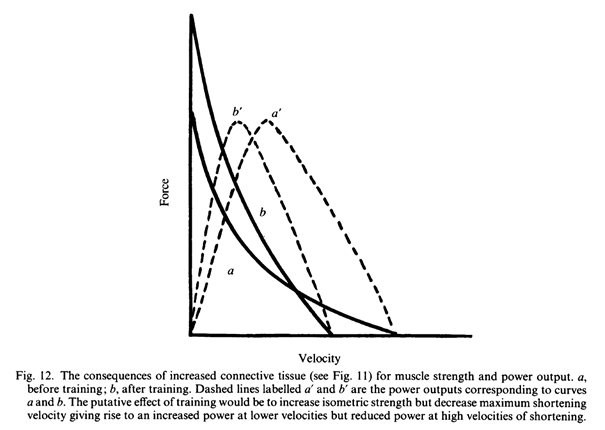
Изменения в строении мышц или соединительной ткани, окружающей мышечные волокна, которые приводят к большей изометрической силе, будут также уменьшать эффективную длину мышечных волокон. Изменение может привести к абсолютному уменьшению длины волокон с изменением углов перистости или, там где соединительная ткань располагаются вдоль волокна (например, рис. 11) возможно уменьшение функциональной длины мышц. В любом случае, в результате уменьшения длины можно было бы ожидать снижения максимальной скорости сокращения мышц, поскольку было бы меньше саркомеров вдоль линии действия мышцы. Сочетание повышенной изометрической силы и снижения скорости сокращения приведут к периодическим изменениям выходной мощности в зависимости от скорости, при которой были сделаны измерения. При малых скоростях можно было бы ожидать, что тренированные мышцы должны быть более мощными, хотя в процентном отношении увеличение будет не таким большим, как исключительно для изометрической силы, в то время как при более высоких скоростях сокращения тренированные мышцы будет менее мощными (рис. 12).
Можно было бы ожидать, что скорость, при которой была достигнута максимальная выходная мощность, уменьшится при снижении эффективной длины мышц. Существует мало экспериментальных данных для проверки этих идей, но многие из экспериментов, касающихся специфики скорости тренировок, как правило, показывают больший прирост в силе при тестировании на низких скоростях (Lesmes et al., 1978;. Caiozzo et al. и рис. 7).
Как выяснилось во время тестирования испытуемых на велоэргометре, тренировка четырехглавой мышцы бедра, результатом которой стал значительный прирост изометрической силы, не привела к увеличению выходной мощности (Rutherford et al. 1986a). Тем не менее, в исследовании приводящей мышцы большого пальца руки, Duchateau & Hainaut (1984) обнаружили, что максимальная скорость сокращения тренированной мышцы не изменилась. Следует отметить, что это исследование было проведено только на одном человеке в пяти точках, чтобы определить кривую зависимости величины силы от скорости.
Уже много было написано о ценности ведения протоколов различных тренировок, и очевидна важность установления наиболее эффективных способов увеличения силы (Atha,1981; McDonagh & Davies, 1984). К сожалению, во многих исследованиях трудно отделить последствия тренировок и возможные нервные адаптации от увеличения силы и гипертрофии отдельных мышц. Тем не менее, есть общее мнение, что должны быть приложены большие усилия, прежде чем будет получен новый прирост мышц. Пока не ясно, является ли это большее усилие само по себе стимулом для изменения или просто это средство обеспечения того, чтобы все ДЕ рекрутировались и тренировались.
Любой рост требует реконструкции существующих структур, то есть увеличения метаболизма, как синтеза, так и деградации. Однако увеличение размера тканей предполагает преобладание белкового синтеза над деградацией белка (Waterlow, Garlick & Millward, 1978).Есть два аспекта метаболизма белка: один из них – это поддержание основного размера ткани. С этой целью метаболизм должен реагировать на краткосрочные изменения, такие как циркуляция аминокислот и глюкозы, а также таких гормонов, как инсулин. Контроль на этом уровне представляет собой, вероятно, контроль на трансляционном уровне с колебаниями в «активности РНК». Второй аспект контроля осуществляется в течение более длительного периода времени. Запасание большего количества ткани, вероятно, зависит от увеличения содержания рибосом, которое, в свою очередь, может потребовать деления клеток-сателлитов и включение одного из дочерних ядер в мышечные волокна. Таким образом, количество ядерного материала возрастает, а размер мионуклеарного домена остается постоянным.
Возможные стимулы для мышечной гипертрофии можно разделить на три категории:
1.Гормональные стимулы
Физическое упражнение может влиять на реакции эндокринной, паракринной и аутокринной систем, что стимулирует мышечный рост. К сожалению, эндокринные изменения являются большим стимулятором, поскольку гипертрофия всегда ограничена на одной группе мышц в одной части тела, что нежелательно. Тем не менее, определенная концентрация, допустимая норма гормонов, таких как инсулин, гормон роста или тестостерон, может быть допустимой, работая в связке с локальными изменениями, связанными с рабочими мышцами. Локальные изменения в паракринных гормонах, таких как инсулиноподобный фактор роста -1 (IGF-1) могут являться очень важными в регуляции роста тканей, но пока не определено, есть ли механизм, объединяющий производство факторов роста и механической активностью мышцы.
2.Метаболические стимулы
Большинство людей тренируется с весом инстинктивно, по принципу «если болит, значит хорошо». Боль (в мышцах) проявляется в жжении, связанном с метаболическими изменениями в работающих мышцах. Несмотря на это общее чувство, здесь нет четкого указания относительно того, что метаболическое истощение является предварительным условием для изменения. Силовые упражнения могут привести к большим метаболическим потокам в тканях с накоплением высоких концентраций H+, неорганического фосфата и креатина, вкупе с небольшими количествами других субстанций, таких как AДФ (аденозиндифосфат), NH3 и инозин, что позволяет одному или нескольким из этих метаболитов стимулировать мышечный рост. Однако изменения этих метаболитов связаны с утомлением, большим, чем от силовых (упражнений усиленной мощности) упражнений и предположительно, что они могут выступать стимулом для быстрого увеличения числа митохондрий и роста капилляров, связанных с повышением выносливости в большей степени, чем просто повышение размера мышц и силы. В тех случаях, когда мы можем наблюдать замедление поступления кислорода к мышцам, и метаболиты могут быть в состоянии хронического истощения, к примеру, при периферических сосудистых заболеваниях, мышечные адаптации приводят к увеличению количества митохондрий в большей степени, чем увеличивается размер волокон. Высоко тренированные на выносливость атлеты не имеют особо сильных мышц.
Физические упражнения, в которых мышечные волокна растягиваются (эксцентрические упражнения или отрицательная работа) могут генерировать гораздо большие силы в активных мышцах, чем обычные упражнения, в которых мышечные волокна укорачиваются, поднимая вес. К тому же, при выполнении эксцентрических упражнений метаболические затраты гораздо меньше, чем при выполнении работы в концентрическом режиме (Abbot, Bigland & Ritchie, 1952). Сравнение тренировок в эксцентрическом (мышца растягивается) и концентрическом (длина мышцы уменьшается) режиме мышечного сокращения позволяет определить средства тестирования. А именно ответить на вопрос: «Что является стимулом мышечного роста: высокая сила, развиваемая мышцей или метаболические изменения?». Мы имеем сведения, что эксцентрические упражнения являются лучшими тренировочными стимуляторами (Komi & Buskirk, 1972), но большинство исследований, использующих этот подход, не показывают разницы между типами упражнений (Rasch, 1974; Jones & Rutherford, 1987), и сравнительная важность силы и метаболических изменений остается до конца неразрешенной.
3.Механические факторы
Хорошо известно, что мышцы находясь растянутом или укороченном состоянии быстро изменяют свою длину таким образом, что длина саркомеров восстанавливается и достигается оптимум силы (Goldspink et al. 1974; Williams & Goldspink, 1978). Это достигается посредством добавления или удаления саркомеров на концах мышечных волокон. В дополнение к изменению длины, мышцы также, кажется, в ответ на растяжения увеличиваются в размерах (Laurent, Sparrow & Millward, 1978).
Есть несколько способов, в которых метаболизм мышц может быть изменен посредством механических воздействий
(1) Большое напряжение может привести к микро-повреждениям структуры саркомера, и тем самым обеспечить стимулы для ремонта и компенсационного роста. Goldspink (1971) предположил, что большое напряжение мышцы приводит к нарушению Z-дисков в результате чего миофибриллы делятся на части, затем они отрастают до полного размера. Это привлекательная визуальная модель и искривление Z линий можно увидеть на электронных микрофотографиях особенно после тренировки с применением эксцентричных сокращений высокой силы (Friden, Sjöstrom & Ekblom, 1983; Newham, McPhail, Mills & Edwards, 1983).
(2) Использование изолированных препаратов диафрагмы крысы или скелетных мышц кролика, Reeds & Palmer (см обзор Reeds, Palmer & Wahle, 1987) показали, что механическая стимуляция (при повторном растяжении покоящейся мышцы) вызывает увеличение как синтеза, так и деградации белков. Авторы также убедительно показали, что это связано с увеличением производства простагландинов, PGF2, будучи связаны с повышенным синтезом и PGE2 с деградации, которая находится в соответствии с результатами Rodermann & Goldberg (1982). Рабочая гипотеза заключается в том, что механическое напряжение вызывает активацию фосфолипазы и освобождение арахидоновой кислоты из мышечных мембран, которые затем служат в качестве прекурсора для синтеза простагландинов.
(3) Коллаген матрицы из соединительной ткани, сформированной фибробластами является неотъемлемой частью мышцы, и она должна расти и меняться вместе с мышечными волокнами. Матрица соединительной ткани и обеспечивает связь между компонентами генерирующими силу и сухожилиями и, следовательно, будет подлежать механическим воздействиям. иРНК для IGF1 были локализованы с фибробластами плода мышцы (Han, D’Ercole & Lund, 1987) и наличие IGF1 обнаружено при регенерации мышечных волокон и клеток-сателлитов после травмы (Jennische, Skottner & Hansson, 1987). Эти наблюдения свидетельствуют о возможном действии паракринной системы, IGF1 синтезируется в фибробластах, но оказывает анаболический эффект на мышечные волокна. Если фибробласты в соединительной ткани продуцируют IGF1 в ответ на механические воздействиям, это может являться способом регулирования роста мышц.
Есть две проблемы с механизмами, описанными выше: во-первых, все они подразумевают некоторую форму повреждения, вероятно, под воздействием больших сил, но как было показано, эксцентрические упражнения не приводят к росту силы (Jones & Rutherford, 1987). Во-вторых, они, вероятно, связаны с изменениями в мышечном волокне, т.е. влияют на метаболизм белка, но не влияют на механизм, посредством которого в мышечное волокно могут поступать дополнительные ядра. Атрофированные мышцы, как результат повреждения или иммобилизации, вероятно, обладают большим потенциалом для роста увеличения силы, чем нормальные неповрежденные мышцы (IMMS, Hackett, Prestidge & Fox, 1977), тем не менее, восстановление, часто является неполным при отсутствии специальной силовой подготовки (Imms & MacDonald, 1978; Imms, 1980). Относительно медленное восстановление контрастирует с быстрым восстановлением силы после экспериментального повреждения мышц. В последнем случае поврежденные волокна дегенерируют, а затем восстанавливаются с помощью клеток-сателлитов до зрелых мышечных волокон за 3-4 недели. У новых волокон, кажется, есть большой потенциал для роста, поскольку повседневная деятельность, как представляется, является достаточным стимулом для полного восстановления силы (Jones, Newham, Round & Tolfree, 1986; Newham, Jones & Clarkson, 1988). Во время восстановления от атрофии, вызванной иммобилизацией, происходит увеличение зрелых волокон, и это может потребовать больше стимулов, для того, чтобы они вернулись к своему первоначальному размеру.
ЗАКЛЮЧЕНИЕ
Тренировочный процесс может быть разделен на несколько этапов. На первом этапе происходит быстрое улучшение способности выполнять силовые упражнения. Это связано с процессом обучения, в котором правильная последовательность мышечных сокращений заложена в качестве моторного паттерна в центральной нервной системе. На этом этапе гипертрофия мышц очень незначительная или вообще отсутствует. Отдельные мышцы не увеличивают силу. Процесс тренировки является весьма специфичным в том, что силовые тренировки развивают силовые способности, но не скоростно-силовые. Вторая фаза увеличения силы отдельных мышц происходит без соответствующего увеличения анатомического поперечника. Механизм этого не ясен, но можно предположить, что это является результатом повышенной нервной активности или какого-то изменения в окружении мышечного волокна или соединительной ткани. Третий этап начинается тогда, когда научные исследования, как правило, заканчиваются (то есть через 12 недель тренировки). В этот момент нетренированные испытуемые выполняют повторную тренировку и тестирование. После этого момента, если тренировка продолжается, то, происходит медленное, но неуклонное увеличение, как размера, так и силы мышц. Стимул этих изменений остается пока загадкой, но почти наверняка он связан с высоким уровнем силы, развиваемой мышцей, что, возможно, вызывает некоторые повреждения, что способствует активации клеток-сателлитов и их включение в существующие мышечные волокна. Наша информация о влиянии долгосрочного обучения в первую очередь поступает из наблюдений на элитных спортсменах, чьи физические показатели вполне могут быть результатом генетического фактора, или в результате использования или злоупотребления анаболических стероидов. Для спортсмена или пациента желающего увеличить размер мышц посредством силовой тренировки наилучшее сочетание интенсивности, частоты и типа упражнения по-прежнему остается вопросом индивидуального выбора, а не научных исследований.
ЛИТЕРАТУРА
- ABBOT, B.C., BIGLAND, B. & RITCHIE, J. M. (1952). The physiological cost of negative work. Journal of Physiology 117, 380-390.
- ALEXANDER, R. McN. & VERNON, A. (1975). The dimensions of the knee and ankle muscles and the forces they exert. Journal of Human Movement Studies 1,115-123.
- ATHA, J. (1981). Strengthening muscle. Exercise and Sports Science Review 9, 1-73.
- BARANY, M. & CLOSE, R.I. (1971). The transformation of myosin in cross-innervated rat muscles. Journal of Physiology 213, 455-474.
- BELANGER, A. Y. & MCCOMAS, A. J. (1981). Extent of motor unit activation during effort. Journal of Applied Physiology 51(5), 1131-1135.
- BELLEMARE, F. & BIGLAND-RITCHIE, B. (1984). Assessment of human diaphragm strength and activation using phrenic nerve stimulation. Respiratory Physiology 58 (3), 263-277.
- BELLEMARE, F., WOODS, J. J., JOHANSSON, R. BIGLAND-RITCHIE, B. (1983). Motor unit discharge rates in maximal voluntary contractions of three human muscles. Neurophysiology 50 (6), 1380-1392.
- BIGLAND-RITCHIE, B. & WOODS, J. J. (1976). Integrated electromyograms and oxygen uptake during positive and negative work. Journal of Physiology 260, 267-277.
- BONDE-PETERSEN, F. (1960). Muscle training by static, concentric and eccentric contractions. Acta physiologica scandinavica 48, 406-416.
- BURKE, R. E. & TSAIRIS, P. (1973). Anatomy and innervation ratios in motor units of cat gastrocnemius Journal of Physiology 234, 749-765.
- CAIOZZO, V. J., PERRINE, J. R. & EDGERTON, V. R. (1981). Training induced alterations of the in vivo force-velocity relationship of human muscle. Journal of Applied Physiology 51 (3), 750-754.
- CERRETELLI, P., MINETTI, A.E., NARICI, M., Roi, G. S. & SCALMANI, B. (1989). Changes in NMR assessed cross-sectional area, force and iEMG activity of human quadriceps during strength training and detraining. Journal of Physiology (in the Press).
- CHAPMAN, S. J., GRINDROD, S. R. & JONES, D. A. (1984). Cross-sectional area and force production of the quadriceps muscle. Journal of Physiology 353, 53P.
- CLARKSON, P. M., KROLL, W. & MCBRIDE, T. C. (1980). Maximal isometric strength and fiber type composition in power and endurance athletes. European Journal of Applied Physiology 44, 35-42.
- CLOSE, R.I. (1972). Dynamic properties of mammalian skeletal muscles. Physiological Review 52, 129-197.
- COYLE,E. F., FEIRING, D. C., ROTKIS, T. C., CoTE III, R. W., ROBY, F. B., LEE, W. & WILMORE, J. H. (1981). Specificity of power improvements through slow and fast isokinetic training. Journal of Applied Physiology 51 (6), 1437-1442.
- CURTIN, N. & DAVIES, R.E. (1972). Chemical and mechanical changes during stretching of activated frog skeletal muscle. Cold Spring Harbor Symposium on Quantitative Biology 37, 619-626.
- DARCUS, H. D. & SALTER, N. (1955). The effect of repeated muscular exertion on muscle strength. Journal of Physiology 129, 325-336.
- DAVIES, C. T. M. & YOUNG, K. (1983). Effects of training at 30% and 100% maximal isometric force (MVC) on the contractile properties of the triceps surae in man. Journal of Physiology 336, 22-23P.
- DAVIES, J., PARKER, D. F., RUTHERFORD, 0. M. & JONES, D. A. (1988). Changes in strength and cross sectional area as a result of isometric strength training. European Journal of Applied Physiology 57, 667-670.
- DELORME, T. L. (1946). Heavy resistance exercises. Archives of Physical Medicine 27, 607-630.
- DONS, B., BOLLERUP, K., BONDE-PETERSEN, F. & HANCKE, S. (1979). The effect of weight lifting exercise related to muscle fibre composition and muscle cross sectional area in humans. European Journal of Applied Physiology 40 (2), 95-106.
- DOOLEY, P., MCDONAGH, M. J. N. & WHITE, M. J. (1983). Training using involuntary electrically evoked contractions does not increase strength. Journal of Physiology 346, 61 P.
- DUCHATEAU, J. & HAINAUT, K. (1984). Isometric or dynamic training: differential effects on mechanical properties of a human muscle. Journal of Applied Physiology 56, 296-301.
- EDGERTON, V. R., Roy, R. R. & APOR, P. (1986). Specific tension of human elbow flexor muscles. In Biochemistry of Exercise VI, p. 487-500. International Series of Sport Sciences, vol. 16, ed. SALTIN, B. Champaign, IL, USA: Human Kinetics Publishers.
- EDJTEHARDI, G. D. & LEWIS, D. M. (1979). Histochemical reactions of fibres in a fast twitch muscle of the cat. Journal of Physiology 287, 439-453.
- EDSTRÖM, L. & GRIMBY, L. (1986). Effect of exercise on the motor unit. Muscle and Nerve 9 (2), 104-126.
- EERBEEK, O., KERNELL, D. & VERHEY, B. A. (1984). Effects of fast and slow patterns of tonic long term stimulation on contractile properties of fast muscle in the cat. Journal of Physiology 352, 73-90.
- FRIDEN, J., SJSTROM, M. & EKBLOM, B., (1983). Myofibrillar damage following intense eccentric exercise in man. International Journal of Sports Medicine 4, 170-176.
- FRONTERA, W. R., MEREDITH, C. N., O’REILLY, K. P., KNUTTGEN, H. G. & EVANS, W. J. (1988). Stength conditioning in older men: skeletal muscle hypertrophy and improved function. Journal of Applied Physiology 64 (3), 1038-1044.
- GANDEVIA, S. C. & MCKENZIE, D. K. (1985). Activation of human diaphragm during maximal static exercise. Journal of Physiology 367, 45-56.
- GARDNER, G. W. (1962). Specificity of strength changes of the exercised and non-exercised limb following isometric training. Respiration Quarterly 34 (1), 98-101.
- GOLDBERG, A. L., ETLINGER, J. D., GOLDSPINK, D. F. & JABLECKI, C. (1975). Mechanisms of workinduced hypertrophy of skeletal muscle. Medicine and Science in Sports and Exercise 7, 248-261.
- GOLDSPINK, G. (1971). Ultrastructural changes in striated muscle fibres during contraction and growth with particular reference to the mechanism of myofibril splitting. Journal of Cell Science 9, 123-138.
- GOLDSPINK, G., TARBRAY, J. C., TARBRAY, C., TARDIEU, C. & TARDIEU, G. (1974). Effect of denervation on the adaptation of sarcomere number and muscle excitability to the functional length of the muscle. Journal of Physiology 236, 733-742.
- GOLLNICK, P. D., TIMSON, B. F., MOORE, R. L. & RIEDY, M. (1981). Muscular enlargement and number of fibres in skeletal muscle of rats. Journal of Applied Physiology 50, 936-943.
- GORDON, E. E., KOWALSKI, K. & FRITTS, M. (1967). Adaptations of muscle to various exercises. Journal of the American Medical Association 199, 103-108.
- GRINDROD, S., ROUND, J. M. & RUTHERFORD, 0. M. (1987). Type 2 fibre composition and force per unit cross sectional area in the human quadriceps. Journal of Physiology 390, 154P.
- GRINDROD, S., TOFTS, P. & EDWARDS, R. H. T. (1983). Investigation of human skeletal muscle structure and composition by X-Ray Computerised Tomography. European Journal of Clinical Investigation 13, 465-468.
- HAN, V. K. M., D’ERCOLE, J. & LUND, P. K. (1987). Cellular localization of somatomedin (insulinlike growth factor) messenger RNA in the human fetus. Science 236, 193-197.
- HELANDER, E. A. S. (1961). Influence of exercise and restricted activity on the protein composition of skeletal muscle. Biochemical Journal 78, 478-482.
- HELLEBRANDT, F. A. & HOUTZ, S. J. (1956). Mechanisms of muscle training in man: Experimental demonstration of the overload principle. Physical Therapy Review 36, 371-383.
- HERZOG, W. & TER KEURS, E. D. J. (1988a). A method for the determination of the force-length relation of selected in-vivo human skeletal muscles. Pflugers Archiv 41, 637-641.
- HERZOG, W. & TER KEURS, E. D. J. (1988b). Force-length relation of in-vivo human rectus femoris muscles. Pfliugers Archiv 411, 642-647.
- HETTINGER, T. (1961). Physiology of Strength. Springfield: C. C. Thomas.
- HORBER, F. F., SCHEIDEGGER, J. T., GRUNIG, B. E. & FREY, F. J. (1985). Thigh muscle mass and function in patients treated with glucocorticoids. European Journal of Clinical Investigation 15, 302-307.
- IKAI, M. & FUKUNAGA, T. (1970). A study on training effect on strength per unit cross sectional area of muscle by means of ultrasonic measurement. Internationale Zeitschrift fur angewandte Physiologie 28, 173-180.
- IMMS, F. J. (1980). The use of physiological techniques for monitoring of progress during rehabilitation following fractures of the lower limb. International Journal of Rehabilitation Medicine 2, 181-188.
- IMMS, F. J., HACKETT, A. J., PRESTIDGE, S. P. & Fox, R. H. (1977). Voluntary isometric strength of patients undergoing rehabilitation following fractures of the lower limb. Rheumatological Rehabilitation 16, 162-171.
- IMMS, F. J. & MAcDONALD, I. C. (1978). Abnormalities of gait occurring during recovery from fracture of the lower leg and their improvement during rehabilitation Scandinavian Journal of Rehabilitation Medicine 10, 193-199.
- JENNISCHE, E., SKOTTNER, A. & HANSSON, H.-A. (1987). Satellite cells express the trophic factor IGF1 in regenerating skeletal muscle. Acta physiologica scandinavica 129, 9-50.
- A. JONES, 0. M. RUTHERFORD AND D. F. PARKER JONES, D. A., NEWHAM, D. J., ROUND, J. M. & TOLFREE, S. E. J. (1986). Experimental human muscle damage: morphological changes in relation to other indices of damage. Journal of Physiology 375, 435-448.
- JONES, D. A. & RUTHERFORD, 0. M. (1987). Human muscle strength training: the effects of three different training regimes and the nature of the resultant changes. Journal of Physiology 391, 1-11.
- KANEHISA, H. & MIYASHITA, M. (1983). Specificity of velocity in strength training. European Journal of Applied Physiology 52, 104-106.
- KEAN, C. J. C., LEWIs, D. M. & MCGARRICK, J. D. (1974). Dynamic properties of denervated fast and slow twitch muscles of the cat. Journal of Physiology 237, 103-113.
- KOMI, P. V. (1986). How important is neural drive for strength and power development in human skeletal muscle? In Biochemistry of Exercise VI, pp. 515-530. International Series on Sport Sciences, vol. 16, ed. SALTIN, B. Champaign, IL, USA: Human Kinetics Publishers.
- KOMI, P. V. & BUSKIRK, E. R. (1972). Effect of eccentric and concentric muscle conditioning on tension and electrical activity of human muscle. Ergonomics 15 (4), 417-434.
- KOMI, P. V., RUSKO, H., VAS, J. & VIHKO, V. (1977). Anaerobic performance capacity in athletes. Acta physiologica scandinavica 100, 107-114.
- KOMI, P. V., VIITASALO, J., RAURAMARA, R. & VIHKO, V. (1978). Effect of isometric strength training on mechanical, electrical and metabolic aspects of muscle function. European Jouranl of Applied Physiology 40, 45-55.
- KRUSE, R. D. & MATHEWS, D. K. (1958). Bilateral effects of unilateral exercise: experimental study based on 120 subjects. Archives of Physical Medicine and Rehabilitation 39, 371-376.
- LARSSON, L. (1982). Physical training effects on muscle morphology in sedentary males at different ages. Medicine and Science in Sports and Exercise 14, 203-206.
- LAURENT, G. J., SPARROW, M. P. & MILLWARD, D. J. (1978). Turnover of muscle protein in the fowl. Changes in rates of protein synthesis and breakdown during hypertrophy of the anterior and posterior latissimus dorsi muscles. Biochemical Journal 176, 407-417.
- LESMES, G. R., COSTILL, D. L., COYLE, E. F. & FINK, W. J. (1978). Muscle strength and power changes during maximal isokinetic training. Medicine and Science in Sports and Exercise 10 (4), 266-269.
- LIND, A. R. & PETROFSKY, J. S. (1978). Isometric tension from rotary stimulation of fast and slow cat muscles. Muscle and Nerve 1, 213-218.
- LINDH, M. (1979). Increase of muscle strength from isometric quadriceps exercise at different knee angles. Scandinavian Journal of Rehabilitation Medicine 11, 33-36.
- LUYTHI, J. M., HOWALD, H., CLAASSEN, H., R6SLER, K., VOCK, P. & HOPPELER, H. (1986). Structural changes in skeletal muscle tissue with heavy resistance exercise. International Journal of Sports Medicine 7, 123-127.
- MCCULLOUGH, P., MAUGHAN, R. J., WATSON, J. S. & WEIR, J. (1984). Biomechanical analysis of the knee in relation to measured quadriceps strength and cross sectional area in man Journal of Physiology 346, 60P.
- MCDONAGH, M. J. N. & DAVIES, C. T. M. (1984). Adaptive response of mammalian skeletal muscle to exercise with high loads. European Journal of Applied Physiology 52, 139-155.
- MCDONAGH, M. J. N., HAYWARD, C. M. & DAVIES, C. T. M. (1983). Isometric training in human elbow flexor muscles: the effects on voluntary and electrically evoked forces. Journal of Bone and Joint Surgery 65, 355-358.
- MACDOUGALL, J. D. (1986). Adaptability of muscle to strength training — a cellular approach. In Biochemistry of Exercise VI, pp. 501-514. International Series on Sport Sciences, vol. 16, ed. SALTIN, B. Champaign, IL, USA: Human Kinetics Publishers.
- MACDOUGALL, J. D., SALE, D. G., MOROZ, J. R., ELDER, G. C. B., SUTTON, J. R. & HOWALD, H. (1979). Mitochondrial volume density in human skeletal muscle following heavy resistance training. Medical Science Sports 11 (2), 164-166.
- MACDOUGALL, J. D., ELDER, G. C. B., SALE, D. G., MOROZ, J. R. & SUTTON, J. R. (1980). The effects of strength training and immobilisation on human muscle fibres European Journal of Applied Physiology 43, 25-34.
- MACDOUGALL, J. D., SALE, D. G., ELDER, G. C. B. & SUTTON, J. R. (1982). Muscle ultrastructural characteristics of elite powerlifters and bodybuilders. European Journal of Applied Physiology 48, 117-126.
- MAUGHAN, R. J. & NIMMO, M. A. (1984). The influence of variations in muscle fibre composition on muscle strength and cross sectional area in untrained males Journal of Physiology 351, 299-311.
- MENDEZ, J. & KEYS, A. (1960). Density and composition of mammalian muscle. Metabolism 9, 184-188.
- MERTON, P. A. (1954). Voluntary strength and fatigue. Journal of Physiology 123, 553-564.
- MILNER-BROWN, H. S., STEIN, R. B. & YEMM, R. (1973). The orderly recruitment of human motor units during voluntary contraction. Journal of Physiology 230, 359-370.
- MOFFROID, M. T. & WHIPPLE, R. H. (1970). Specificity of speed of exercise. Physical Therapy 50, 1693-1699.
- MORITANI, T. & DEVRIES, H. A. (1979). Neural factors versus hypertrophy in the time course of muscle strength gain. American Journal of Physical Medicine 58, 115-130.
- NEWHAM, D. J., JONES, D. A. & CLARKSON, P. M. (1988). Repeated high-force eccentric exercise:
- effects on muscle pain and damage. Journal of Applied Physiology 63, 1381-1386.
- NEWHAM, D. J., MCPHAIL, G., MILLS, K. R. & EDWARDS, R. H. T. (1983). Ultrastructural changes after concentric and eccentric contractions. Journal of Neurological Science 61, 109-122.
- PENMAN, P. A. (1970). Human striated muscle ultrastructural changes accompanying increased strength without hypertrophy. Respiration Quarterly 41, 418-424.
- RACK, P. M. & WESTBURY, D. R. (1969). The effects of length and stimulus rate on tension in the isometric cat soleus muscle. Journal of Physiology 204, 443-460.
- RASCH, P. J. (1974). The present status of negative exercise — A review. American Corrective Therapy Journal 28 (3), 77-94.
- RASCH, P. J. & MOREHOUSE, L. E. (1957). Effect of static and dynamic exercises on muscular strength and hypertrophy. Journal of Applied Physiology 11 (1), 29-34.
- RASCH, P. J., PIERSON, W. R. (1964). One versus multiple positions in isometric exercise. American Journal of Physical Medicine 43, 10-12.
- RASCH, P. J., PIERSON, W. R. & LOGAN, G. A. (1961). The effect of isometric exercise upon the strength of antagonistic muscles. Internationale Zeitschrift fur angewandte Physiologie 19, 18-22.
- REEDS, P. J. & PALMER, R. M. (1985). The role of prostaglandins in the control of muscle protein turnover. In Control and Manipulation of Animal Growth, ed. BUTTERY, P. J., LINDSAY, D. B. & HAYNES, N. B., pp. 161-185. London: Butterworths.
- REEDS, P. J., PALMER, R. M. & WAHLE, K. W. J. (1987). The role of metabolites of arachidonic acid in the physiology and pathophysiology of muscle protein metabolism. Biochemical Society Transactions 15, 328-331.
- RODEMANN, H. P. & GOLDBERG, A. L. (1982). Arachidonic acid, prostaglandin E2 and F2X influence rates of protein turnover in skeletal and cardiac muscle Journal of Biological Chemistry 257 (4), 1632-1638.
- ROYCE, J. (1964). Re-evaluation of isometric training methods, A Must. Research Quarterly 35, 215-216.
- RUTHERFORD, 0. M. (1986). The determinants of human muscle strength and the effects of different high resistance training regimes. Ph.D. Thesis, University College London.
- RUTHERFORD, 0. M. (1988). Muscular coordination and strength training: implications for injury rehabilitation. Sports Medicine 5, 196-202.
- RUTHERFORD, 0. M., GREIG, C. A., SARGEANT, A. J. & JONES, D. A. (1986a). Strength training and power output: transference effects in the human quadriceps muscle. Journal of Sports Science 4, 101-107.
- RUTHERFORD, 0. M. & JONES, D. A. (1988). Contractile properties and fatiguability of the human adductor pollicis and first dorsal interosseous: a comparison of the effects of two chronic stimulation patterns. Journal of Neurological Science 85, 319-331.
- RUTHERFORD, 0. M. & JONES, D. A. (1986). The role of learning and coordination in strength training. European Journal of Applied Physiology 55, 100-105.
- RUTHERFORD, 0. M., JONES, D. A. & NEWHAM, D. J. (1986b). Clinical and experimental application of the percutaneous twitch superimposition technique for the study of human muscle activation. Journal of Neurology and Neurosurgical Psychiatry 49, 1288-1291.
- SALE, D. G. & MACDOUGALL, J. D. (1981). Specificity in strength training; a review for the coach and athlete. Canadian Journal of Applied Sports Science 6, 87-92.
- SALMONS, S. & HENRIKSSON, K. (1981). The adaptive response of skeletal muscle to increased use. Muscle and Nerve 4, 94-105.
- SALMONS, S. & VRBOVA, G. (1969). The influence of activity on some contractile characteristics of mammalian fast and slow muscles Journal of Physiology 201, 535-549.
- SARGEANT, A. J., HOINVILLE, E. & YOUNG, A. (1981). Maximum leg force and power output during short term dynamic exercise. Journal of Applied Physiology 51 (5), 1175-1182.
- SCHIAFFINO, S., BORNIOLI, S. P. & ALIOSI, N. (1972). Cell proliferation in rat skeletal muscle during early stages of compensatory hypertrophy. Virchows Archiv Abteilung B. Zellpathologie 11, 268-273.
- STREET, S. F. (1983). Lateral transmission of tension in frog myofibres: a myofibrillar network and transverse cytoskeletal connections are possible transmitters. Journal of Cellular Physiology 114, 346-364.
- TESCH, P. & KARLSSON, J. (1978). Isometric strength performance and muscle fibre type distribution in man. Acta physiologica scandinavica 103, 47-51.
- TESCH, P. & KARLSSON, J. (1982). Muscle hypertrophy in bodybuilders. European Journal of Applied Physiology 49, 301-306.
- THEPAUT-MATHIEU, C., HOECKE, J. & MATON, B. (1988). Myoelectrical and mechanical changes linked to length specificity during isometric training. Journal of Applied Physiology 64 (4), 1500-1505.
- THOMAS, D. O., WHITE, M. J., SAGAR, G. & DAVIES, C. T. M. (1978). Electrically evoked isokinetic plantar flexor torque in males. Journal of Applied Physiology 63, 1499-1503.
- THORSTENSSON, A., GRIMBY, G. & KARLSSON, J. (1976a). Force-velocity and fibre composition in human knee extensor muscle. Journal of Applied Physiology 40, 12-16.
- THORSTENSSON, A., KARLSSON, J., VIITASALO, J. H. T., LUHTANEN, P. & KOMI, P. V. (1976 b). Effect of strength training on EMG of human skeletal muscle. Acta physiologica scandinavica 98, 232-236.
- WATERLOW, J. C., GARLICK, P. J. & MILLWARD, D. J. (1978). Protein Turnover in Mammalian Tissues and in the Whole Body. Amsterdam: North Holland.
- WILKIE, D. R. (1950). The relation between force and velocity in human muscle. Journal of Physiology 110, 249-280.
- WILLIAMS, P. & GOLDSPINK, G. (1978). Changes in sarcomere length and physiological properties in immobilized muscle. Journal of Anatomy 127, 459-468.
- YOUNG, A. (1984). The relative isometric strength of Type 1 and Type 2 muscle fibres in the human quadriceps. Clinical Physiology 4, 23-32.
- YOUNG, A., STOKES, M., ROUND, J. M. & EDWARDS, R. H. T. (1983). The effect of high resistence training on the strength and cross-sectional area of the human quadriceps. European Journal of Clinical Investigation 13, 411-417.
Перевод выполнили магистранты: Н.А. Балякина, А.Ю. Баранова, Ю.А. Гарамов, С.А. Красношлык, А.Д. Фомичева, А.С. Кудрявцева, М.В. Мелихова, А.В. Панин, Н.В. Судакова, Е.В. Тарасова, К.В. Цебро, А.В. Александрова, А.В. Миллер, Г.А. Попов, З.В. Фролова, Н.П. Микуляк.
Редакция перевода – А.В. Самсонова
1989_Jones-et-al_perevod.pdf