Эксцентрические сокращения вызывают повреждение мышечных волокон
Доказано, что эксцентрические сокращения вызывают повреждение мышечных волокон. Сильные повреждения наблюдаются в сократительном и цитоскелетном компонентах мышечного волокна. Степень повреждения мышцы после эксцентрических упражнений зависит от архитектуры мышц, типа мышечных волокон, а также от степени напряжения мышцы.
Frieden J, Lieber R.L. Eccentric exercise-induced injuries to contractile and cytoskeletal muscle fibre components //Acta Physiol. Scand. 2001, 171, P.321-326.
Фрайден Ж, Либер Р.Л.
ЭКСЦЕНТРИЧЕСКИЕ СОКРАЩЕНИЯ ВЫЗЫВАЮТ ПОВРЕЖДЕНИЯ В СОКРАТИТЕЛЬНОМ И ЦИТОСКЕЛЕТНОМ КОМПОНЕНТАХ МЫШЕЧНОГО ВОЛОКНА
1 Department of Hand Surgery, Sahlgrenska University Hospital, GoEteborg, Sweden 2 Departments of Orthopaedics and Bioengineering, University of California and Veterans Affairs, San Diego, USA
РЕЗЮМЕ
При упражнении удлиняющаяся мышца может быть повреждена. Недавние сообщения документировали механизм повреждения, вызванного упражнениями, а также физиологические и клеточные события, происходящие при этом. В начале тяжелых эксцентрических упражнений имеет место потеря цитоскелетного белка десмина и потеря клеточной целостности, как свидетельство повреждения сарколеммы. Эти исследования указывают, что самые ранние события в повреждении мышцы являются механическими по природе, в то время как более поздние события указывают на то, что эксцентрические упражнения могут активизировать процесс регенерации мышцы. Мы считаем, что степень повреждения мышцы после эксцентрического упражнения различна в мышцах с различной архитектурой, зависит от типа мышечных волокон, а также от степени напряжения мышечных волокон в эксцентрической фазе и усилена воспалением после начального повреждения.
Получена 12 декабря 2000, принята 24 января 2001
DOMS – явление мышечного дискомфорта и боли в течение активных сокращений, которые достигают максимума через 24 – 48 часов после напряженного упражнения. В начале боль локализуется в проксимальных и дистальных отделах мышцы, затем распространяется на всю мышцу. Боль в мышце и ее повреждение связаны с интенсивностью работы, особенно если она для человека непривычна. Однако, даже люди в отличной спортивной форме могут испытывать боли в мышцах, и их повреждения, если работа для них новая. Хотя падение уровня силы наблюдается очень быстро (в течение минут) после выполнения упражнения, основные причины отношения между DOMS и потерей силы мышцы еще недостаточно ясны.
ЭКСЦЕНТРИЧЕСКИЙ РЕЖИМ МЫШЕЧНОЙ АКТИВНОСТИ
Erling Asmussen еще в 1950 году установил, что DOMS непосредственно связаны с эксцентричным компонентом упражнения (Asmussen 1952, 1956). В течение эксцентрического действия, сокращающаяся мышца вынужденно удлиняется. В течение концентрического действия мышца продуцирует напряжение, укорачиваясь. При изометрическом сокращении мышца напряжена при неизменной длине. Все три действия — общие компоненты ежедневного движения. Напряжение, генерируемое при эксцентрическом сокращении больше, чем при концентрическом и изометрическом (рис.1) (Katz 1939, Singh и 1966 Karpovich, Komi и Раско 1974, Stauber 1989), хотя ДЕ активировано меньше (Bigland-Ritchie и Woods, 1976).
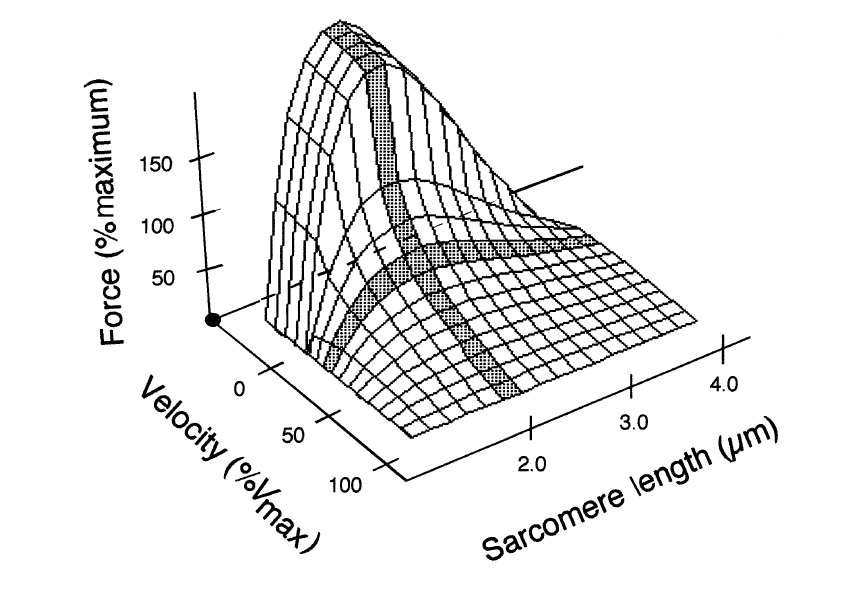
В дополнение к относительным различиям в напряжении между режимами мышечного сокращения, рис. 1 также демонстрирует изменения в напряжении, как функции скорости сокращения. При концентрическом режиме отношение напряжение/скорость описывается уравнением Хилла (Hill, 1938) и связано с взаимодействием актина и миозина (Huxley 1969). При эксцентрическом режиме напряжение имеет две фазы. Сначала, мышца сокращается, продуцируя силу, затем мышца пассивно растягивается внешней силой, хотя генерирует дополнительное напряжение (Katz 1939, Stauber 1989). Пассивное растяжение побудило исследователей подозревать, что упругий компонент неповрежденной системы «сухожилие – мышца» производит дополнительное напряжение (см. обзор Chapman, 1985). Недавние исследования предполагают, что пассивное напряжение мышцы определяет специфическая изоформа титина и что она различается у различных скелетных мышц, даже в пределах тех же самых типов (Wang и др. 1993, Labeit и 1995 Kolmerer). Другие исследователи считают, что энергетика формирования поперечного мостика в течение фазы сокращения/растяжения является причиной более высокого напряжения для эксцентрической деятельности (Stauber 1989). Недавно, Linari и сотрудники демонстрировали, что при удлинении число поперечных мостиков увеличивается, хотя в среднем степень напряжения поперечных мостиков подобна вызванному изометрическому сокращению (Linari и др. 2000).
ОБЪЕКТИВНЫЕ ДАННЫЕ, СВЯЗАННЫЕ С DOMS
Основные данные, связанные DOMS включают: понижение мышечной силы, уменьшение подвижности и увеличенный уровень креатинкиназы в сыворотке крови (CK). Множество исследований демонстрировало, что эксцентрические сокращения приводят к увеличению уровня креатинкиназы в течение 24–48 часов после упражнения, который может сохраниться в течение 3–6 дней. Креатинкиназа — внутримышечный фермент, ответственный за то, чтобы поддержать адекватные уровни ATФ в течение мышечного сокращения. Его появление в крови интерпретируется как увеличенная проницаемость или разрушение мембраны мышечного волокна. Восприятие боли после эксцентрического упражнения не совпадает с уровнем CK в сыворотке крови (Newham и др. 1988). Также важно отметить, что существенные повреждения мышцы и их некроз, демонстрируемый в экспериментах на животных, не могут быть воспроизведены при выполнении эксцентрических упражнений людьми. Исследователи демонстрировали защитный эффект предыдущей тренировки, сравнивая уровни CK с уровнем у нетренированных объектов из контрольной группы (Evans и др. 1985). Часто предполагается, что уровень CK так или иначе связан с величиной повреждения мышцы, хотя эта потенциальная корреляция не была явно доказана. В предварительном исследовании 26 кроликов, выполняющих эксцентрические движения посредством m. dorsi. exors мы нашли слабую корреляцию между уровнем CK в сыворотке крови и понижением силы мышцы, осуществляющей движения (рис. 2) (Friden и Lieber, 2000).
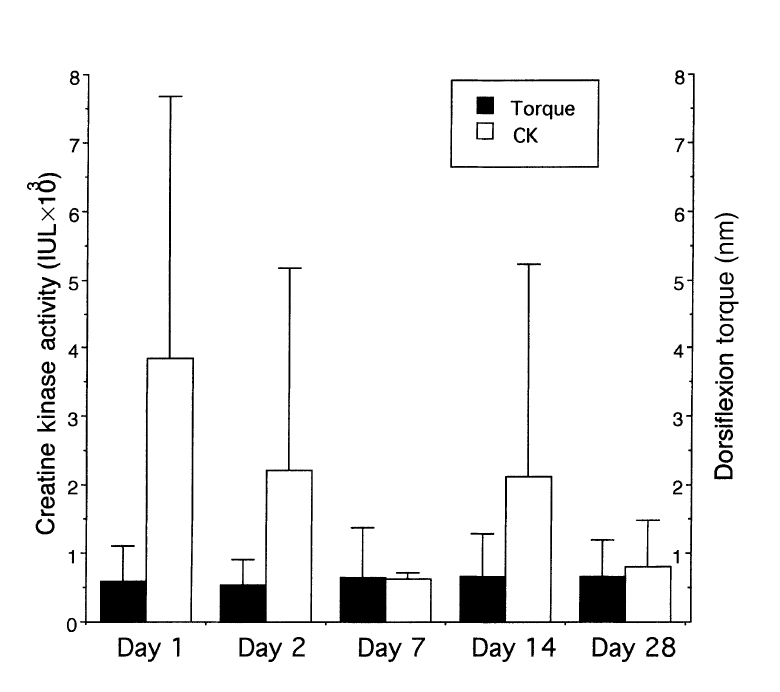
Возможно, что эти отношения являются относительно слабыми, потому что проницаемость мышечного волокна для внутримышечных ферментов может или не может быть связана с его сокращением. Например, мы предварительно демонстрировали, что многочисленные мышечные волокна, подвергшиеся эксцентрическому сокращению, которые сохранили их способность исключить плазматический фибронектин, демонстрировали существенные структурные отклонения, типа потери внутриклеточного десмина, разрушение миофибрилл и разрушение Z-дисков.
ПОВРЕЖДЕНИЕ МЫШЦ
Непосредственное уменьшение силы после эксцентрического сокращения связано как с растяжением саркомера, что приводит к неоптимальному наложению актина и миозина (Faulkner и др. 1993), так и с изменениями в их сцеплении (Balnave и Аллен 1995, Morgan и Аллен 1999). Faulkner и сотрудники предложили, что в течение эксцентрического упражнения некоторые саркомеры сильно растянуты, и таким образом повреждены, в то время как другие сохраняют свою длину. Известно, что эксцентрическое упражнение вызывает большие изменения, чем сопоставимые сокращения при изометрической или концентрической деятельности (Warren и др. 1993). Несмотря на большую потерю силы после эксцентрических сокращений, мембранный потенциал покоя изолированных мышц, изнуренных эксцентрическими или изометрическими сокращениями был одинаков (Warren и др. 1993).
Поскольку нет никаких свидетельств в отказе возбуждения, способность производить потенциалы действия, как ожидается, будет незатронутой. Независимо от точного механизма, найдено соглашение, что начальное снижение силы вызвано механическим повреждением. Множество механических факторов, типа длины мышцы, силы и скорости, кажется, играют роль в последствиях эксцентрических сокращений. Newham и др. (1988) нашел, что длинные мышцы теряют больше силы после эксцентрических сокращений по сравнению с короткими. Хотя морфологические повреждения мышцы хорошо документированы, механизм повреждения до сих пор полностью не понят. Хотя ткань мышцы является чрезвычайно пластичной (Pette 1990), разрушительные изменения ультраструктуры мышцы могут быть ответом на необычные условия (Hoppeler 1986). Имеются сообщения о морфологических отклонениях после различных типов упражнений в экспериментах на животных (Armsrtong и др. 1983, Kuipers и др. 1983, McCully и в 1985 Faulkner, Lieber и 1988 Friden) и упражнениях, выполняемых людьми (Friden и др. 1981, 1983, 1984, Newham и др. 1983). Различные типы и местоположения клеточных повреждений после тяжелого упражнений показаны в таблице 1.
Таблица 1 Доказательства изменений в мышце, связанные с упражнениями
| Величина мышечного повреждения | Автор |
| 1.Первичое или вторичное повреждение сарколеммы | Jenkins, 1988, Armsrtong, 1990, Duan et all, 1990 |
| 2. набухание или разрушение саркотубулярной системы. | Friden, Lieber, 1996, Armsrtong 1990, Willems et all. 1999 |
| 3 Искажение сократительных компонентов миофибрилл | Armsrtong et all. 1983, Newham et all. 1983, Friden et all. 1984, Friden et all . 1988) |
| 4 Повреждение цитоскелета | Friden et all. 1984, Lieber et all 1996, Friden, Lieber ,1998 |
| 5. Повреждения внеклеточной миофибриллярной матрицы | Stauber, 1989 |
Огромное количество морфологических исследований указывает, на то, что Z-диск — самая уязвимая структура в вызванном повреждении мышцы посредством эксцентрических упражнений. Повреждение было также найдено в сарколемме, T-трубочках, миофибриллах и цитоскелете. При умеренном типе повреждения, сразу после окончания упражнения повреждается немного саркомеров. Обычно наблюдается «вытекание» Z-дисков (появление волн) с нарушением порядка миофибрилл. В более серьезных случаях, может произойти обширное вытекание (появление волн и расширение Z-дисков) и «размазывания» Z-диска (дисперсия Z-дискового материала в соседний саркомер); потеря центровки Z-диском и смещения Z-дискового материала. Повреждаются волокна I типa и II типа, хотя большинство исследований демонстрирует большие повреждения в волокнах II типа (Lieber и Friden, 1988). Показано, что белки саркоплазматического ретикулума могут играть существенную роль в этиологии повреждения мышцы (Gilchrist и др. 1992). Несколько исследований указывают, что нарушение метаболизма кальция может быть связано с изменениями мышцы и ее «слабостью» после тяжелых упражнений (Reid и др. 1994). В последних исследованиях выдвинута гипотеза, что вначале изменения происходят с цитоскелете (Lieber и др. 1996), а именно в цитоскелетных белках, расположенных на уровне Z-диска (α-актинин, виментин и десмин). Самое поразительное изменение, наблюдаемое после эксцентрического упражнения в мышцах dorsi.exion кролика – потеря десмина, расположенного в значительной части мышечных волокон поперек мышцы (Lieber и др. 1996). Три дня после эксцентрических упражнений, более чем 30 % волокон EDL потеряли окрашивание десмина. Этот процент быстро уменьшился, так, что после 7 дней процент был приблизительно 10 % для EDL. После 28 дней, менее 1% мышечных волокон обнаружили повреждения десмина. Интересно, что большинство, но не все мышечные волокна с разрушенным десмином (десмин-отрицательные волокна) были также фибронектин положительны (то есть имели неразрушенный фибронектин), что указывает на потерю целостности мышечного волокна, сопровождающей разрушение цитоскелета. Нужно отметить, что несмотря на то, что десмин был разрушен и удален из мышечного волокна (отсутствовало окрашивание десмина), мышечное волокно все еще сохраняло сократительные и окислительные ферменты. Belcastro (1993) сообщил об увеличении деятельности калпаинов в мышцах нижних конечностей после бега на тредмилле. Он постулировал, что ранние механические события, которые приводят к повреждениям, а затем к воспалению, требуют наличия промежуточных событий. Наиболее вероятные промежуточные события – увеличение внутриклеточного уровня кальция (Balnave и Аллен 1995). В то время как циклические изменения концентрации кальция нормальны в цикле сокращения мышцы, хроническое повышение уровня кальция может активировать эндогенные протеазы (например, калпаины), вызывая ухудшение в работе мышцы. Как результат, увеличение внутриклеточного кальция остается привлекательной особенностью большинства моделей повреждения мышцы. Belcastro также демонстрировал увеличенную сходство калпаина для кальция после упражнения, предлагая, что упражнение увеличивает суммарное содержание калпаинов, чтобы разрушить ткань мышцы, как и сходство для кальция. Данное количество повреждения могло таким образом произойти после упражнения при более низкой концентрации кальция. Интересно, что калпаин – специфичен для белков цитоскелета, тогда как актин и миозин, кажется, не затронуты. Два белка, связанные с промежуточной системой филаментов (α–актинин и десмин) сильно повреждены.
МОРФОЛОГИЧЕСКИЕ ПРОЯВЛЕНИЯ
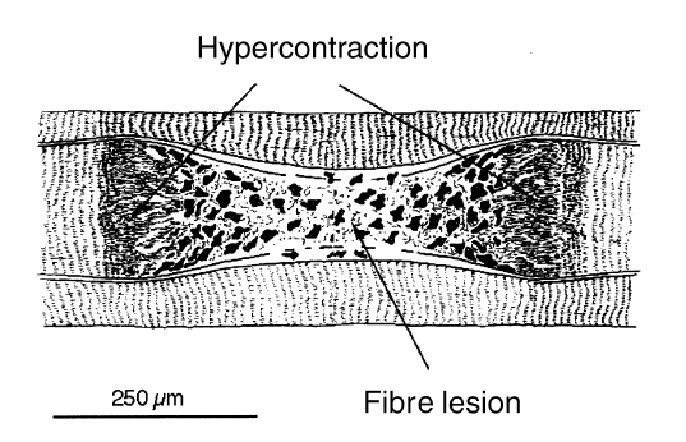
В многочисленных предыдущих исследованиях повреждения мышц, выполненных на животных, мышечные волокна с неправильными формами и размерами использовались как морфологические показатели острого, вызванного упражнением повреждения мышцы (Armsrtong и др. 1983, McCully и Faulkner 1985, Lieber и Friden 1988, Friden и др. 1991, Lieber и др. 1991). В результате эксцентрических упражнений были продемонстрированы очень большие волокна. Эти волокна были идентифицированы, как быстрые гликолитические (FG). Кроме того, в этих больших волокнах часто демонстрировалось уменьшение саркоплазменного фибронектина, что свидетельствовало о повреждении мембраны мышечного волокна (Friden и др. 1991). Большие волокна, которые были найдены в гистологических исследованиях после повторных эксцентрических сокращений, представляют частичное гиперсокращение волокна (Friden и Lieber,1998). Этот феномен имеет место при повреждении мембраны мышечного волокна и его некрозе, что проявляется в очень коротких саркомерах (рис. 3).
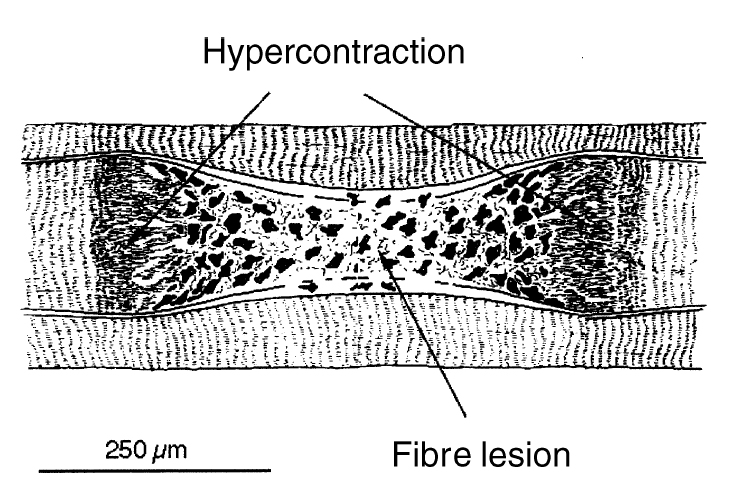
С обеих сторон повреждения, волокно восстановило свои сократительные элементы, и миофибриллы стали все более и более плотно упакованными. Большие миофибриллы появляются на различных расстояниях от «выпуклых зон» как миофибриллы с нормальной толщиной или значительно более широкие, чем соседние «нормальные» миофибриллы. В пограничном районе между широкими и тонкими частями волокна, были найдены клетки-сателлиты, хотя их консистенция была значительно меньшей, чем в тонкой области мышечного волокна. В тонкой области, мембрана клетки очень редко оставалась неповрежденной и через три дня после эксцентрических упражнений в ней присутствовали многочисленные макрофаги. Важно помнить, что эти «чрезвычайные» модели упражнения, возможно, непосредственно не касаются моделей DOMS у человека, где демонстрировалось намного меньше повреждений. После эксцентрических упражнений у кролика, ультраструктурные исследования показали, что типичная решетка A-полосы была последовательно потеряна, и Z-дисковые решетки, и даже Z-дисковый материал отсутствовал в биопсиях, взятых через один час после упражнения (Friden и Lieber 1996). Тонкие и толстые нити были сильно дезорганизованы, и остатки Z-дисков были волнистыми и поврежденными во многих местах. Саркоплазматический ретикулум и система Т-трубочек в биопсиях, взятых через час после упражнения были или нормальны или расширены, в то время как в биопсиях, взятых значительно позже эксцентрического упражнения они казались округленными и увеличенным, а также имеющими гранулярную плотность. Волокна с невысокой плотностью десмина, но отрицательным окрашиванием фибронектина, по-видимому соответствуют непосредственному ультраструктурным последствиям под воздействием эксцентрического груза о которых предварительно было сообщено, то есть, искажение выравнивания A и I дисков, нерегулярные Z-диски и уменьшение толстых нитей из тонкой решетки нити (Дикс и Расселл 1991, Lieber и др. 1991). Разумно предположить, что целостная система волокна находится по существу в хорошем структурном состоянии, если центральные повреждения не появляются многократно в одном и том же самом волокне. Это, однако, не возможно, потому что, как только волокно разрушено, на оставшимися неповрежденными волокна, возможно, подает большее напряжение. Это означает, что в конечном итоге меньше волокон будет подвергнуто принудительному удлинению, как был предложено ранее (Friden и др. 1983, Armsrtong 1990). С другой стороны, центрально поврежденное волокно может функционировать как обычное, особенно если рассмотреть факт, что продольные промежуточные нити могут охватывать травмированные доли и передать напряжение (Wang и др. 1993).
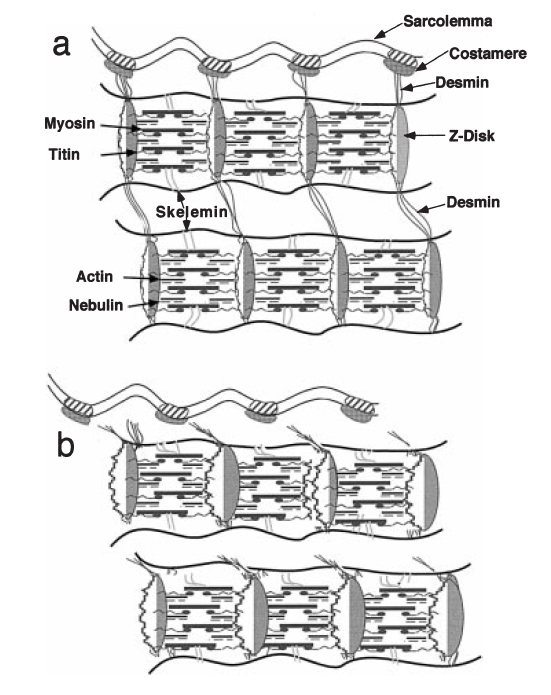
Чтобы проверить роль десмина в нормальной и травмированной мышце, мышцы EDL были получены от молодых диких мышей, и гомозиготных мышей, у которых отсутствовал ген, отвечающий за формирование десмина (Milner и др. 1996, Сэм и др. 2000). Мышцы dorsi.exors выполняли эксцентрические упражнения на эргометре, который мог осуществлять быстрые изменения длины. Были обнаружены достоверные различия в проценте повреждений Z-дисков в экспериментальной и контрольной мышце, но не было никаких достоверных различий у мышей, у которых отсутствовал ген, отвечающий за десмин и дикими мышами. С другой стороны, была найдена достоверно более высокая степень бокового смещения миофибрилл у мышей, у которых отсутствовал ген, контролирующий производство десмина. Эти данные демонстрируют различия между Z-дисковым разрушением и Z-дисковым смещением. Увеличенное продольное смещение Z-дисков вдоль продольной оси миофибрилл, свидетельствует о том, что у мышей-нокаутов (без гена десмина) Z-диски менее хорошо «заякорены» относительно друг друга (рис. 4).
АДАПТАЦИЯ
Может быть неправильно говорить, что упражнение приводит к повреждению мышцы. Лучше сказать, что интенсивные эксцентрические упражнению запускают процесс ремоделирования мышцы, который приводит к гипертрофии. Существенно, что декремент силы связан с эксцентрическими упражнениями и не связан с изометрическими при той же длительности. Таким образом, мы выдвигаем гипотезу, что эксцентричное упражнение запускает события, которые приводят к насыщению мышцы лейкоцитами и моноцитами из кровотока и к проникновению этих клеток в мышечную ткань. Этот клеточный инфильтрат приводит к дальнейшей деградации ткани, вероятно смещением протеолитических ферментов, необходимых для регенерации ткани. Исследование эффекта нестероидных наркотиков (NSAIDs) после эксцентрического упражнения, продемонстрировало, что введение урбипрофена после повреждения мышцы может улучшить сократительную функцию и уменьшить циркулирующие уровни воспаления клеток и ферментов мышцы (Mishra и др. 1995). Эти данные предлагают, что процесс воспаления является вторичным, и также приводит к DOMS. Морфологический механизм быстрой адаптации, вероятно, заключается в том, чтобы сделать миофибриллы, цитоскелет и клеточную мембрану более стойкими к напряжению и стрессу. (Gibala и др. 1995). Быстрое восстановление демонстрировалось после эксцентрического упражнения quadriceps. Hortobagyi и др. (1998) нашел, что это восстановление произошло независимо от разрушения клетки и заключил, что это связано с нервными факторами. Результаты Hortobagyi были основаны на факте, что после второй эксцентрической тренировки ЭМГ, амплитуда сухожильного рефлекса и уровень CK были в норме, хотя 23 % ткани мускула были повреждены. В заключении мы выдвигаем гипотезу о порядке событий при выполнении упражнений, после которых наблюдается DOMS: разрушение цитоскелета, что влечет смещение миофибрилл в мышечном волокне, повреждение Z-дисков и дезорганизацию A-полосы, потерю целостности мышечного волокна как проявление состояния сарколазменного фибронектина, гиперсокращение травмированных областей волокна и вторжение в волокно воспалительных клеток. Профилактические и терапевтические действия могут действовать на несколько из этих направлений, но до сих пор нет консенсуса по этому вопросу.
ЛИТЕРАТУРА
- Armstrong, R.B. 1990. Initial events in exercise-induced muscular injury. Med Sci Sports Exerc 22, 429±435.
- Armstrong, R.B., Ogilvie, R.W. & Schwane, J.A. 1983. Eccentric exercise-induced injury to rat skeletal muscle. J Appl Physiol 54, 90±93.
- Asmussen, E. 1952. Positive and negative muscular work. Acta Physiol Scand 28, 364±382.
- Asmussen, E. 1956. Observations on experimental muscular soreness. Acta Rheum Scand 2, 109±116.
- Balnave, C.D. & Allen, D.G. 1995. Intracellular calcium and force in single mouse muscle fibres following repeated contractions with stretch. J Physiol (Lond) 488, 25±36.
- Belcastro, A. 1993. Skeletal muscle calcium-activated neutral protease (calpain) with exercise. J Appl Physiol 74, 1381±1386.
- Bigland-Ritchie, B. & Woods, J.J. 1976. Integrated electromyogram and oxygen uptake during positive and negative work. J Physiol (Lond) 260, 267±277.
- Chapman, A.E. 1985. The mechanical properties of human muscle. Ex Sport Sci Rev 13, 443±501.
- Dix, D.J. & Russell, B.R. 1991. Redistribution of heavy myosin chain mRNA in the midregion of stretched muscle fibers. Cell Tissue Res 263, 61±69.
- Duan, C., Delp, M.D., Hayes, D.A., Delp, P.D. & Armstrong, R.B. 1990. Rat skeletal muscle mitochondrial [Ca2+] and injury from downhill walking. J Appl Physiol 68, 1241±1251.
- Evans, W.J., Meredith, C.N., Cannon, J.G. et al. 1985. Metabolic changes following eccentric exercise in trained and untrained men. J Appl Physiol 61, 1864±1868.
- Faulkner, J.A., Brooks, S.V. & Opiteck, J.A. 1993. Injury to skeletal muscle fibres during contractions: conditions of occurrence and prevention. Phys Ther 73, 911±921
- Friden, J. & Lieber, R.L. 1996. Ultrastructural evidence for loss of calcium homeostasis in exercised skeletal muscle. Acta Physiol Scand 158, 381±382.
- J Friden and R L Lieber _ Exercise-induced muscle injury Friden, J. & Lieber, R.L. 1998. Segmental muscle fiber lesions after repetitive eccentric contractions. Cell Tissue Res 293, 165±171.
- Friden, J. & Lieber, R.L. 2001. Serum creatine kinase level is a poor predictor of muscle force after injury. Scand J Med Sci Sports 11, 126±127
- Friden, J., SjoEstroEm, M. & Ekblom, B. 1981. A morphological study of delayed muscle soreness. Experientia 37, 506±507.
- Friden, J., SjoEstroEm, M. & Ekblom, B. 1983. Myo fibrillar damage following intense eccentric exercise in man. Int J Sports Med 4, 170±176.
- Friden, J., KjoE rell, U. & Thornell, L.E. 1984. Delayed muscle soreness and cytoskeletal alterations. An immunocytological study in man. Int J Sports Med 5, 15±18.
- Friden, J., Seger, J. & Ekblom, B. 1988. Sublethal muscle fiber injuries after high-tension anaerobic exercise. Eur J Appl Physiol 57, 360±368.
- Friden, J., Lieber, R.L. & Thornell, L.E. 1991. Subtle indications of muscle damage following eccentric contractions. Acta Physiol Scand 142, 523±524.
- Gibala, M.J., MacDougall, J.D., Tarnopolsky, M.A. & Stauber, W.T. 1995. Changes in human skeletal muscle ultrastructure and force production after acute resistance exercise. J Appl Physiol 78, 702±708.
- Gilchrist, J., Wang, K., Katz, S. & Belcastro, A.N. 1992. Calcium-activated neutral protease effects upon skeletal muscle sarcoplasmic reticulum protein structure and calcium release. J Biol Chem 267, 20857±20865.
- Hill, A.V. 1938. The heat of shortening and the dynamic constants of muscle. Proc Roy Soc Lond Series B: Biol Sci (London) 126, 136±195.
- Hoppeler, H. 1986. Exercise-induced ultrastructural changes in skeletal muscle. Int J Sports Med 7, 76±92.
- Hortobagyi, T., Houmard, J., Fraser, D., Dudek, R., Lambert, J. & Tracy, J. 1998. Normal forces and myo fibrillar disruption after repeated eccentric exercise. J Appl Physiol 84, 492±498.
- Huxley, H.E. 1969. The mechanism of muscular contraction. Science 164, 1356±1366.
- Jenkins, R.R. 1988. Free radical chemistry. Sports Med 5, 156±170.
- Katz, B. 1939. The relation between force and speed in muscular contraction. J Physiol (Lond) 96, 45±64.
- Komi, P.V. & Rusko, H. 1974. Quantitative evaluation of mechanical and electrical changes during fatigue loading of eccentric and concentric work. Scand J Rehabil Med 3, 121±126.
- Kuipers, H., Drukker, J., Frederik, P.M., Guerten, P. & Kranenburg, G.V. 1983. Muscle degeneration after exercise in rats. Int J Sports Med 4, 45±51.
- Labeit, S. & Kolmerer, B. 1995. Titins: giant proteins in charge of muscle ultrastructure and elasticity. Science 270, 293±296.
- Lieber, R.L. & Friden, J. 1988. Selective damage of fast glycolytic muscle fibers with eccentric contraction of the rabbit tibialis anterior. Acta Physiol Scand 133, 587±588.
- Lieber, R.L., McKee-Woodburn, T. & Friden, J. 1991. Muscle damage induced by eccentric contractions of 25% strain. J Appl Physiol 70, 2498±2507.
- Lieber, R.L., Thornell, L.-E. & Friden, J. 1996. Muscle cytoskeletal disruption occurs within the first 15 minutes of cyclic eccentric contraction. J Appl Physiol 80, 278±284.
- Linari, M., Lucii, L., Reconditi, M. et al. 2000. A combined mechanical and X-ray diffraction study of stretch potentiation in single frog muscle fibres. J Physiol (Lond) 526, 589±596.
- McCully, K.K. & Faulkner, J.A. 1985. Injury to skeletal muscle fibers of mice following lengthening contractions. J Appl Physiol 59, 119±126.
- Milner, D.J., Weitzer, G., Tran, D., Bradley, A. & Capetanaki, Y. 1996. Disruption of muscle architecture and myocardial degeneration in mice lacking desmin. J Cell Biol 134, 1255± 1270.
- Mishra, D.K., Friden, J., Schmitz, M.C. & Lieber, R.L. 1995. Antiin.ammatory medication after muscle injury. A treatment resulting in short-term improvement but subsequent loss of muscle function. J Bone Joint Surg (Am) 77, 1510±1519.
- Morgan, D. & Allen, D. 1999. Early events in stretch-induced muscle damage. J Appl Physiol 87, 2007±2015.
- Newham, D.J., McPhail, G., Mills, K.R. & Edwards, R.H.T. 1983. Ultrastructural changes after concentric and eccentric contractions of human muscle. J Neurol Sci 61, 109±122.
- Newham, D.J., Jones, D.A., Ghosh, G. & Aurora, P. 1988. Muscle fatigue and pain after eccentric contractions at long and short length. Clin Sci 74, 553±557.
- Pette, D. 1990. The Dynamic State of Muscle Fibers. Walter de Gruyter, Berlin.
- Reid, W., Huang, J. & Bryson, S. 1994. Diaphragm injury and myo fibrillar structure induced by resistive loading. J Appl Physiol 76, 176±184.
- Sam, M., Shah, S., Friden, J., Milner, D.J., Capetanaki, Y. & Lieber, R.L. 2000. Desmin knockout muscles generate lower stress and are less vulnerable to injury compared with wild-type muscles. Am J Physiol 279, C1116±C1122.
- Singh, M. & Karpovich, P.V. 1966. Isotonic and isomenbsp; tric forces of forearm .exors and extensors. J Appl Physiol 21, 1435±1437.
- Stauber, W.T. 1989. Eccentric action of muscles: physiology, injury, and adaptation. Ex Sport Sci Rev 17, 157±185.
- Wang, K., McCarter, R., Wright, J., Beverly, J. & Ramirez- Mitchell, R. 1993. Viscoelasticity of the sarcomere matrix of skeletal muscles. The titin-myosin composite filament is a dual-stage molecular spring. Biophys J 64, 1161±1177.
- Warren, G.I., Hayes, D.A., Lowe, D.A., Prior, B.M. & Armstrong, R.B. 1993. Mechanical factors in the initiation of eccentric contraction-induced injury in rat soleus muscle. J Physiol (Lond) 464, 457&plu smn;475.
- Willems, M., Huijing, P.A. & Friden, J. 1999. Swelling of sarcoplasmic reticulum in the periphery of muscle fibres after isometric contractions in rat semimembranosus lateralis muscle. Acta Physiol Scand 165, 347±356.
Перевод А.В.Самсоновой