Адаптация различных типов мышечных волокон к нагрузкам
Адаптация различных типов мышечных волокон к нагрузкам представлена в статье Старон Р.С. с соавт. При помощи биопсии определяли композицию мышечных волокон широкой латеральной мышцы бедра трех групп людей: контрольной, тяжелоатлетов, и бегунов. Гистохимический анализ образцов биопсии показал, что у бегунов было высокое процентное содержание МВ I и IIC типа, чем у групп контроля и у тяжелоатлетов.
Staron, R.S. Human Skeletal Muscle Fiber Type Adaptability to Various Workloads / R.S. Staron, R.S. Hikida, F.C. Hagerman, G.A. Dudley, T.F. Murray //The Journal of Histochemistry and Cytochemistry, 1984.– Vol. 32.– No 2.– pp.146-152.
Р.С.СТАРОН С СОАВТ.
АДАПТАЦИЯ РАЗЛИЧНЫХ ТИПОВ МЫШЕЧНЫХ ВОЛОКОН К НАГРУЗКАМ РАЗЛИЧНОЙ НАПРАВЛЕННОСТИ
Кафедра зоологических и биомедицинских наук, Ирвин Холл, Университет Огайо, Афины, штат Огайо 45701
РЕЗЮМЕ
При помощи биопсии взяты образцы из широкой латеральной мышцы бедра трех групп людей: контрольной, тяжелоатлетов, и бегунов. Группа бегунов оказалась уникальной группой по отношению к переменным измерения (низкая масса тела и процент жира в организме, и высокий объем потребления кислорода). Кроме того, гистохимический анализ образцов биопсии показал, что у бегунов было высокое процентное содержание МВ I и IIC типа, чем у групп контроля и у тяжелоатлетов. Использовав криостатический метод поиска, каждое из волокон определено гистохимически, затем анализировано морфометрически с помощью электронного микроскопа. Результаты измерения процентного объема митохондрий показали сильную взаимосвязь между АТФазой и окислительной способностью типов волокон для всех трех групп таким образом, что окислительная активность будет ранжирована I> IIA> IIB. Независимо от типа волокна, существуют значительные различия между группами в отношении количества митохондрий мышечного волокна (бегуны> тяжелоатлеты> контроль) и содержания липидов (бегуны > контроль > тяжелоатлеты). У тяжелоатлетов содержание митохондрий больше, чем у контрольной группы. Можно предположить, что отсутствие активности способствует низкому процентному содержанию митохондрий и высокому проценту мышечных волокон IIB типа.
КЛЮЧЕВЫЕ СЛОВА:
Типы мышечных волокон человека, морфометрический анализ, спортсмены
ВВЕДЕНИЕ
До недавнего времени, ультраструктурные исследования адаптации типов мышечных волокон человека к различным нагрузкам проводились с помощью косвенных методов (Kiessling et al., 1975; Orlander et al., 1977; Prince et al., 1981). Так как скелетные мышцы человека состоят из нескольких типов волокон (Jonson et al., 1973; Edgerton et al., 1975), трудно охарактеризовать эти типы волокон ультраструктурно. Применяя криостатический метод определения состава скелетных мышц человека, различные типы мышечных волокон, смешанные в мышце, могут быть собраны гистохимическим способом, а затем проанализированы морфометрическим методом посредством электронной микроскопии (Ingjer, 1979a; Sjostorm et al., 1982а; Staron et al., 1983). В настоящем исследовании мы использовали эту технику для характеристики и сравнения нормальных типов мышечных волокон у нетренированных лиц, тяжелоатлетов и бегунов, тренированных на выносливость.
МАТЕРИАЛЫ И МЕТОДЫ
Испытуемые
Данные были получены от 20 человек, разделенных на три группы: контрольную (5), тяжелоатлеты (7), и бегуны (8). Нетренированные люди вели чрезвычайно малоподвижный образ жизни, не имея регулярной физической активности. И тяжелоатлеты и бегуны были дисциплинированными спортсменами, тренировались 4-6 дней в неделю в течение не менее 3 лет.
Тяжелоатлеты, (в том числе два тяжелоатлета – призеры соревнований и один бодибилдер высокого уровня) не включали в тренировку аэробные упражнения любого рода. Точно так же, бегуны (имеется в виду время пробегания марафона = 159 мин.) не включали силовую тренировку нижних конечностей в их ежедневную тренировку. Широкая латеральная мышца бедра была выбрана для этого исследования из-за ее доступности, распределения типов волокон, и участие как в тренировке тяжелоатлетов (приседания, жимы ногами, и т.д.), так и у бегунов (бег на возвышенность). Некоторые из этих данных были ранее опубликованы (Dudley et al., 1983; Staron et al., 1983). Антропометрические и соответствующие метаболические данные приведены в таблице 1.
Таблица 1. Физические данные и соответствующие метаболические измерения
| Группы | Возраст, лет | Рост, см | Вес, кг | %ВF* | МПК мл/кг мин |
Контрольная группа | |||||
| 1 | 20 | 178,0 | 82,9 | 18,1 | 46,1 |
| 2 | 38 | 185,4 | 109,8 | 21,0 | — |
| 3 | 36 | 176,5 | 72,5 | 20,3 | 31,6 |
| 4 | 19 | 175,7 | 65,0 | 9,0 | 47,3 |
| 5 | 32 | 184,0 | 86,5 | 25,7 | 40,6 |
| Среднее | 29 | 179,9 | 83,3 | 18,8 | 41,4 |
Тяжелоатлеты | |||||
| 1 | 20 | 181,5 | 84,1 | 11,7 | 45,7 |
| 2 | 21 | 169,0 | 72,6 | 11,7 | 46,9 |
| 3 | 21 | 182,0 | 94,6 | 13,3 | 51,1 |
| 4 | 31 | 170,0 | 86,5 | 10,9 | 44,5 |
| 5 | 21 | 168,0 | 102,0 | 25,5 | 39,3 |
| 6 | 26 | 169,0 | 57,5 | 13,3 | — |
| 7 | 28 | 176,5 | 92,5 | 21,9 | 34,9 |
| Среднее | 24 | 173,7 | 85,7 | 15,5 | 43,7 |
Бегуны | |||||
| 1 | 25 | 175,0 | 62,0 | 10,9 | 65,0 |
| 2 | 26 | 170,5 | 61,3 | 11,3 | 63,1 |
| 3 | 27 | 180,5 | 69,7 | 10,5 | — |
| 4 | 22 | 165,5 | 58,5 | 5,5 | 61,2 |
| 5 | 27 | 173,5 | 70,6 | 6,2 | 59,5 |
| 6 | 37 | 186,7 | 67,6 | 10,1 | 55,4 |
| 7 | 26 | 177,8 | 65,5 | 9,0 | 56,9 |
| 8 | 31 | 163,0 | 60,9 | 11,3 | 56,2 |
| Среднее | 28 | 174,0 | 164,5 + + | 9,4 + | 59,6 + + |
Примечание:+ – значительно отличается (р <0,05). + + – значительно отличается (р <0,01).
% BF – Процент жира
Физические и метаболические данные
Формула Brozek et al., (1963) была использована для вычисления процента жира из расчета удельного веса (через гидростатическое взвешивание) и остаточной емкости легких (Wilmore et al., 1980). Каждый испытуемый выполнял тест на возрастающую нагрузку максимального потребления кислорода (МПК) на велосипедном эргометре фирмы Monank (см. Dudley et al., 1983). Скорость вращения педалей была 60 об/мин при нагрузке 30 Вт на первую минуту. Рабочая нагрузка впоследствии снижалась на 30 Вт в минуту до момента, пока не нарушался ритм. Поглощение кислорода за минуту было определялось полуавтоматической системой (Wilmore и Costill, 1974). МПК определялся как максимальный уровень потребления кислорода с увеличением рабочей нагрузки.
Подготовка образцов
Образцы мышцы были взяты посредством биопсии из поверхностной части латеральной широкой мышцы бедра у 20 исследуемых, используя технику биопсийной иглы Bergstrom (1962) с изменениями, внесенными Evans et al. (1982). Данные о распределении типов волокон в широкой латеральной мышце различаются (Lexell др, 1983;. Nygaard и Sanchez, 1982; Edgerton и др., 1975; Johnson и др., 1973); однако были приняты меры, чтобы наиболее точно определить идентичную локацию для биопсии у всех испытуемых. Повторные биопсии (по крайней мере, одна на группу) показали бессистемные и незначительные изменения в распределении типов волокон. Кроме того, биопсия из икроножной мышцы трех бегунов содержала аналогичное распределение типов волокон, как и в латеральной широкой мышце (Green и др., 1981).
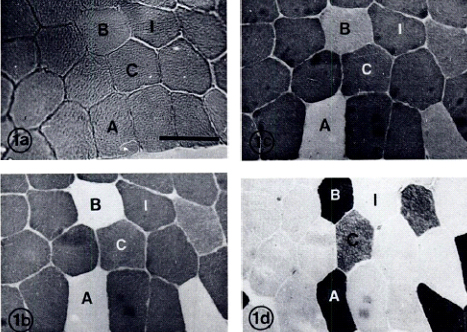
Образцы мышц были помещены в трагакант, и охлаждены в изопентане до — 160ºС жидким азотом. Серийные срезы (10-µм толщиной) были вырезаны на криостате (American Optical Buffalo, NY) при- 20ºС и помещены на стекла для гистохимического анализа. Этот анализ состоял из пробирного анализа для оценки активности миофибриллярной АТФазы при рН 4,3, 4,6, и 10,4 (Brooke и Kaiser, 1970; Staron et al., 1983). Типы мышечных волокон были разделены на четыре группы (типы I, IIA, IIB, IIC) на основании устойчивости или лабильности их АТФазы в преинкубационной среде (Рисунок 1). Тип I (медленно-сокращающиеся) волокна были стабильны в кислотных диапазонах, но лабильны при рН=10,4. Мышечные волокна IIA типа показали обратную картину. Все волокна, которые были стабильными при рН 10,4 и 4,6, но нестабильны при 4,3 рН были классифицированы как Титип IIB. Типы волокна IIAB (см. Staron et al., 1983) были сгруппированы в зависимости от типа IIB волокон. Остальные волокна были классифицированы как IIC, стабильные во всем диапазоне рН (хотя были волокна с различными уровнями интенсивности окрашивания). Значения площади поперечного сечения по меньшей мере 25 волокон на каждый тип волокна мышечного образца определяли используя Keuffel и Essen (Morristown, NJ) компенсационный полярный планиметр, направленный при 200-кратном увеличении. Данные поперечного сечения не были зарегистрированы, если не набиралось 25 волокон каждого типа волокна в образце.
Периодически выполнялись срезы толщиной 40-50 µм и подготовлялись для электронного микроскопа (Lngjer, 1977; Eisenbeng и Kuda, 1977). Эти толстые замороженные срезы сразу помещали в охлажденный (2-4ºС) реактив Карновского в (2% формальдегид и 2,5% глутарового альдегида в 0,1 М фосфатного буфера, 7,1 рН) и фиксировали в течение 2 часов. Затем промывали в фосфатно-буферной сахарозе, фиксировали в OsO4 на час при 4 ºС в обезвоженном этаноле, и передавались для заключения Epon и Analdite. Тонкие образцы были нарезаны ультрамикротом Reichert ОМ U2, закрепленным на медную решетку с прорезями, с поддержкой сьемки, противопоставлены уранилацетату и цитрату свинца и исследованы в электронном микроскопе Zeiss ЕМ 109.
Морфометрический анализ
Мышечные волокна, выявленные гистохимическим анализом, были исследованы с помощью электронно-микроскопических методов (Weibel et al., 1966; Eisenberg and Kuda, 1975). Три-четыре микрофотографии на каждое мышечное волокно были сделаны при увеличении в 8000 раз. Анализ пяти мышечных волокон на определение типа волокна одного физического лица, состоял в общей сложности почти из 300 проанализированных мышечных волокон. Микрофотографии были закодированы, чтобы сохранить анонимность, процентный объем из митохондрии и капель триглицерида были определены в сечениях в середине мышечного волокна (исключая субсарколемные скопления).
Статистический анализ
Двусторонний дисперсионный анализ (ANOVA) был задействован для всех данных. Угловое преобразование было сделано от процентного содержания в объеме митохондрий и капель триглицерида перед выполнением анализа ANOVA этих параметров. Значительные различия между значениями всех переменных были определены с использованием апостериорного теста Student-Newman-Keuls.
РЕЗУЛЬТАТЫ
Согласно различным значениям, бегуны были уникальной группой (низкий вес тела и процент жира в организме, и высокий уровень МПК), Таблица 1. Распределение типов волокн показало более высокий процент МВ I типа и IIC типа у выносливых тренируемых испытуемых по сравнению с тяжелоатлетами и нетренированными лицами (Таблица 2). Четверо человек (трое нетренирующихся и один тяжелоатлет) имели нестандартное распределение типов волокон. Тип IIB волокна составил 64,5% из всех образцов биопсии. Эти данные были получены в предыдущей работе (Staron et al., 1983).
Поперечные сечения трех типов волокон (I, IIA и IIB) были одинаковы для бегунов и нетренирующихся (Таблица 3). Однако волокна IIA типа в мышцах тяжелоатлетов были значительно крупнее, чем у любого нетренирующегося и у бегунов. Они были также крупнее, чем МВ I типа и IIB типа.
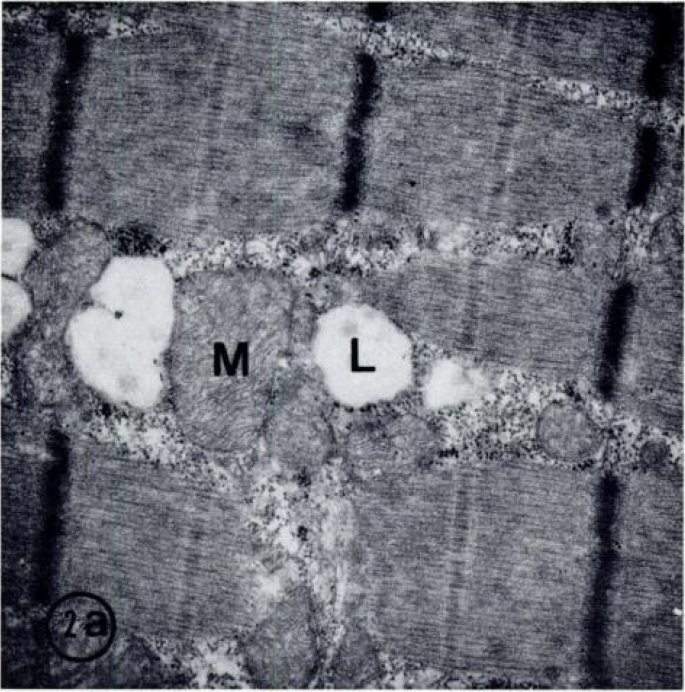
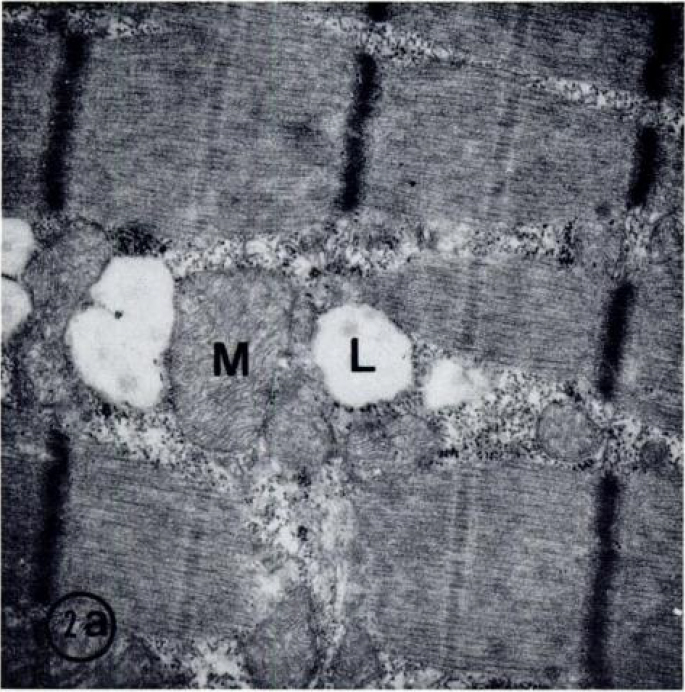
Гистохимически выявленные типы волокон были исследованы ультраструктурно с помощью стереологического метода. С препаративной техникой, которую мы использовали, целостность клетки хорошо сохраняется для ультраструктурного исследования (рис. 2). Митохондрии и липидные капли оказались нормальными. Объемная плотность (%) митохондрий значительно отличается между типами I, IIA, и IIB (I>IIA>IIB) волокон для всех трех групп испытуемых (Таблица 4). Волокна данного типа у бегунов и тяжелоатлетов содержали значительно более высокое количество митохондрий, чем волокна группы контроля. У тяжелоатлетов и бегунов имелось похожее процентное содержание митохондрий в мышечных волокнах IA типа и IIB типа.
Таблица 2 Распределение мышечных волокон различных типов, %
| I | IIA | IIB | IIC | n | |
| Контроль | |||||
| 1 | 34,7 | 49,6 | 15,7 | 0,0 | 427 |
| 2 | 13,4 | 21,8 | 64,8 | 0,0 | 627 |
| 3 | 15,0 | 8,2 | 76,8 | 0,0 | 607 |
| 4 | 23,3 | 76,2 | 0,5 | 0,0 | 206 |
| 5 | 28,8 | 12,8 | 58,4 | 0,0 | 541 |
| Среднее | 23,1(9,0) | 33,7 (28,7) | 43,2 (33,2) | 0,0 (0,0) | 482 |
| Тяжелоатлеты | |||||
| 1 | 31,5 | 42,1 | 25,6 | 0,8 | 387 |
| 2 | 45,6 | 43,5 | 10,4 | 0,5 | 643 |
| 3 | 37,9 | 36,0 | 26,1 | 0,0 | 261 |
| 4 | 59,3 | 40,5 | 0,0 | 0,2 | 830 |
| 5 | 20,7 | 21,4 | 57,7 | 0,2 | 646 |
| 6 | 30,2 | 54,9 | 14,7 | 0,2 | 572 |
| 7 | 42,1 | 39,4 | 18,3 | 0,2 | 837 |
| Средние | 38,2 * (12,5) | 39,7 (10,0) | 21,8 (18,2) | 0,3 (0,3) | 597 |
| Бегуны | |||||
| 1 | 70,5 | 20,8 | 5,3 | 3,4 | 379 |
| 2 | 76,7 | 15,6 | 0,0 | 7,7 | 506 |
| 3 | 59,5 | 21,6 | 11,4 | 7,5 | 914 |
| 4 | 58,0 | 31,4 | 3,0 | 7,6 | 761 |
| 5 | 59,2 | 28,6 | 2,1 | 10,1 | 898 |
| 6 | 77,1 | 22,4 | 0,2 | 0,3 | 1048 |
| 7 | 71,7 | 25,2 | 0,0 | 3,1 | 1000 |
| 8 | 72,7 | 21,3 | 5,7 | 0,2 | 877 |
| Среднее | 68,2+ + (8,0) | 23,4 (4,9) | 3,4 ** (1,9) | 5,0 + + (3,7) | 798 |
Примечание n – количество волокон. В скобках () указано стандартное отклонение. Значительные отличия от группы контроля: * – р <0,05, ** – р = 0,01. Значительно отличается от тяжелоатлетов и группы контроля: + + – Р <0,01.
Содержание липидов было выше (P <0,01) во всех трех типах волокон у бегунов, чем в группе контроля и у тяжелоатлетов, за исключением МВ IIB типа в группе контроля (Таблица 4). Количество липидов значительно отличалось (р <0,05) между тремя различными типами волокон в группе бегунов (I>IIA> IIB), а также между медленно сокращающимися (МВ I типа) и быстро-сокращающимися (МВ IIA и IIB типов) у тяжелоатлетов. Процентный объем липидов в волокнах IIA типа у тяжелоатлетов и в группе контроля также был различным (р <0,05). Независимо от типа волокна, три группы значительно отличались процентным объемом содержания митохондрий (бегуны > тяжелоатлеты> контроль) и процентным объемом липидов (бегуны > контроль > тяжелоатлеты) (таблица 5).
ОБСУЖДЕНИЕ
Для классификации типов мышечных волокон в скелетных мышцах человека были использованы два способа. Первый основан оценке содержания окислительных ферментов, второй – по активности миофибриллярной АТФ-азы (см Saltin et al., 1977). Из-за наложения окислительных ферментативных компонентов между типами волокон, классификации двух систем не совместимы (Sjogaard et al., 1978;. Reichmann и Pette, 1982). Тем не менее, в нашем исследовании была установлена тесная взаимосвязь между активностью АТФазы и окислительной способностью, если волокна рассматривать как группы (I>IIA>IIB) (таблица 4). Это было продемонстрировано биохимически (Essen et al., 1975.); по семиквантитативныму определению содержания митохондрий (Ingjer, 1979a, b); косвенно c помощью структуры М-полосы (Sjostrom и др., 1982b; Angquist и Sjostrom, 1980); и стереологическим способом по гистохимически определенным волокнам (Staron и соавт., 1983). Тем не менее, способ классификации мышечных волокон по активности АТФазы, также может быть упрощением пластичности скелетных мышц млекопитающих. Изменение среды рН предварительной инкубации дает множество подтипов волокон: IIC, IIAC, IIA, IIAB, и IIB. Пожалуй, нецелесообразно использовать такое большое количество типов волокон но, они могут отражать превращения, происходящие в результате количества и вида их вовлечения в мышце.
Группа бегунов в нашем исследовании имела значительный процент содержания МВ I типа (среднее значение = 68,2%), по сравнению с другими двумя группами. Это было хорошо задокументировано для бегунов на длинные дистанции (Prince et al., 1976; Costill et al., 1976; Gollnick et al., 1972). Является ли преобладание МВ I типа у этих выносливых спортсменов индуцируемым посредством тренировок или генетически предопределено – требует дополнительных исследований. В то время как большинство данных поддерживают утверждение не конвертируемости между типами I и II МВ у людей (см. Gollnick, 1982), несколько исследований показали, преобразование между этими двумя типами волокон (см. Howald, 1982).
Таблица 3 Средние значения площади поперечного сечения (стандартная погрешность)
| I | IIA | IIB | IIC | |
| Контроль | 4730 (927) | 6860 (1754)+ | 6184 (2327) | — |
| Тяжелоатлеты | 5290 (647) ++ | 9111 (2137) | 6743 (611) + | — |
| Бегуны | 4943 (1116) | 4555 (1331) ++ | 4378 (1785) | 4868 (1055) |
Примечание: Площадь измеряется мкм². Значительно меньше, чем у тяжелоатлетов МВ IIA типа: + р <0,05, + + р <0,01.
Хотя это не наша инициатива на основе данного исследования поперечного сечения делать выводы о возможном преобразовании между медленными и быстрыми сокращающимися волокнами, но интересно отметить, что у бегунов в исследовании, увеличивался процент МВ IIC типа (5,0%). Обычно, МВ IIC типа составляет 0-2% от общего количества волокон в мышцах человека (Dubowitz and Brooke, 1973). Эти волокна были названы IIC-IIAC (Ingjer, 1979a, б), недифференцированные или неклассифицированные (Saltin et al., 1977; Dubowitz и Brooke, 1973), или промежуточные (Schantz et al., 1982) волокна. Было предположено, что в здоровой мышце выносливых спортсменов, они играют роль во взаимопревращении между типами I и волокон II (Jansson et al., 1978; Schantz et al., 1982).
Существует суждение, что трансформация происходит в быстросокращающихся МВ скелетных мышцах человека, как с интенсивной тренировкой на выносливость (IIB→IIA) (Jansson и Kaijser, 1977; Anderson и Henriksson, 1977; Greenet и др., 1979) и иммобилизации (IIA→IIB) (Haggmark, 1979). Это, казалось бы, объясняет низкий процент IIB волокон у выносливых спортсменов и высокий процент IIB волокон у малоподвижных лиц. Costill et al., (1976) показали, положительное отношение между МПК и процентным содержанием МВ I типа. Точно так же, наше исследование показывает отрицательную корреляцию между процентом МВ II типа и МПК (г = -0,728, Р <0,01).
Объемная плотность митохондрий (%) в мышечных волокнах IIB типа у бегунов и тяжелоатлетов была одинаковой, но значительно выше, чем в контрольной группе. Похоже, что МВ IIB типа могут быть в первую очередь гликолитическими, и после тренировки могут развивать высокую окислительную способность, не нарушая общую иерархию метаболического потенциала (I> IIA> IIB). Тем не менее, должна быть некоторая точка, в которой происходит изменение молекулы миозина в быстро сокращающихся МВ.
Эксперименты на истощение гликогена у людей показали, что рекрутирование МВ IIB типа зависит от интенсивности и продолжительности упражнений (Essen, 1978; Green,1978; Thomson et al., 1979). Мышцы тяжелоатлетов подвергаются высокой интенсивности, почти максимальным сокращениям, и у этой группы спортсменов можно было бы ожидать значительный процент МВ IIB типа. Тем не менее, тяжелоатлеты в нашем исследовании не имеют значительно более высокий процент МВ IIB типа, чем контроль. Кроме того, у пауэрлифтеров, как оказалось, преобладают МВ IIA типа (на 32,8% крупнее, чем те же волокна у группы контроля). Аналогичным образом, Costill и соавт. (1979) обнаружили увеличение площади IIA/I и IIA/IIB типов МВ после силовой тренировки. Вполне может быть, что кумулятивный эффект от силовых упражнений, включает большее вовлечение МВ IIA типа, чем I или IIB типа.
В долгосрочном исследовании, MacDougall et al., (1979) обнаружили корреляцию между снижением процента объемной плотности митохондрий и увеличением площади быстро сокращающихся волокон в тяжелых силовых тренировках.
Процент объемной плотности митохондрий в нашем исследовании был значительно больше у тяжелоатлетов, чем в контрольной группе (сравнивались как конкретные типы волокон, так и все волокна вместе). Это несоответствие может быть связано с использованием косвенного метода (MacDougall et al., 1979) в отличие от прямого (настоящее исследование) морфометрического анализа, или, возможно, с низким уровнем активности группы контроля в нашем исследовании. Тем не менее, МПК в двух группах было одинаково. Как и ожидалось, силовые тренировки были связаны с увеличением активности анаэробных АТФ-генерирующих систем мышцы (Thorstensson et al., 1976; Costill et al., 1979). Силовые тренировки также могут увеличить аэробную производительность АТФ (Costill et al., 1979) и увеличить краткосрочную выносливость без соответственного увеличения МПК (Hickson et al., 1980). Таким образом, может быть, бездействие способствует низкой объемной плотности митохондрий и высокому проценту МВ IIB типа, что наблюдалось у контрольной группы.
Таблица 4 Средние значения морфометрических данных
| I | IIA | IIB | IIC | |
| Контроль | ||||
| Объем митохондрий% | 3,04 (0,94)++** | 2,25 (0,75)++ ** | 1,62 (0,55)++** | |
| Объем липидов% | 0,29 (0,23) | 0,22 (0,23)* | 0,13 (0,16) | |
| n | 25 | 25 | 20 | |
| Тяжелоатлеты | ||||
| Объем митохондрий% | 4,11 (1,13) + + | 3,01 (0,87) ** | 2,21 (0,72) + + | 3,43 (1,80) ** |
| Объем липидов% | 0,24 (0,30) + ** | 0,13 (0,19) ** | 0,05 (0,09) ** | 0,10 (0,12) * |
| n | 35 | 35 | 30 | 4 |
| Бегуны | ||||
| Объем митохондрий% | 4,43 (0,94) + | 3,94 (1,31) | 2,37 (1,19) + + | 4,64 (0,96) + |
| Объем липидов% | 0,82 (0,64) + | 0,53 (0,39) | 0,26 (0,36) + | 0,53 (0,5 1) |
| n | 40 | 39 | 16 | 28 |
Примечание: * – значимые отличия от типа волокна A + = р <0,05. + + = Р <0,01. Значимые отличия от того же типа волокна в нижних группах: * – (р <0,05), ** – (р <0,01). n – количество волокон. В скобках () – стандартное отклонение
Это не удивительно, что объемная плотность липидов (%) у бегунов была значительно больше, чем у тяжелоатлетов и в группе контроля. Запасы триглицерида – это важный внутримышечный субстрат для тренировок на выносливость (Еssen et al., 1977; Lithell et al., 1979). Трудно интерпретировать различия в объемной плотности липидов (%) между тяжелоатлетами и группой контроля. Это может быть просто погрешность выборки, так как распределение липидов в волокне обычно варьируется, и многие из волокон у паэрлифтеров содержали 0% липидов (как определено в соответствии с нашими морфометрическими методами).
В заключение хотим сказать, что эти данные демонстрируют важную ультраструктурную взаимосвязь между окислительной способностью (содержание митохондрий и липидов) и типом мышечного волокна человека (I> IIA > IIB), что сохраняется с различными схемами тренировок. Процентное содержание митохондрии внутри МВ типов I, IIA, и IIB было значительно больше у тяжелоатлетов и бегунов, по сравнению с нетренированными людьми. Таким образом, силовые упражнения (которые в значительной степени могут опираться на МВ IIA типа) могут способствовать увеличению окислительной способности мышечных волокон без существенного влияния на МПК. В отличие от этого, в волокнах мышц контрольной группы было отмечено низкое процентное содержание митохондрий во всех трех типах волокон и высокий процент МВ IIB типа, вероятно, в результате низкого уровня активности этой группы.
Таблица 5 Групповые итоги для морфометрических данных *
| Показатель | Контроль | Тяжелоатлеты | Бегуны |
| Объем митохондрий% | 2,35 (0,96) | 3,16 (1,22) | 4,06 (1,30) |
| Объем липидов% | 0,22 (0,22) | 0,14 (0,23) | 0,59 (0,54) |
| n | 70 | 104 | 123 |
*Все данные значимо отличаются друг от друга (P <0,01).
n = количество волокон. () = Стандартное отклонение.
Благодарности
Авторы благодарят Mike Wigal за техническую помощь в сборе физических и метаболических данных, и доктора Jon’a Wairo и Joseph’a Steinmetz за их помощь в статистических анализах. Особая благодарность тем людям, которые были волонтерами для этого исследования.
ЛИТЕРАТУРА
- Anderson P, Henniksson J (1977): Training-induced changes in thesub groups of human type II skeletal muscle fibres. Acta Physiol Scand 99: 123
- Angquist K-A, Sjostrom ( 1980): Intermittent claudication and muscle fiber fine structure: morphometnic data on mitochondrial volumes. Ultrastruct Pathol 1:461
- Bergstrom J (1962): Muscle electrolytes in man. Scand J Clin Lab Invest 14(Suppl)68: 1
- Brooke MH, Kaiser KK (1970): Three “myosin ATPase” systems: the nature of their pH lability and sulfydry dependence. J Histochem Cytochem 18:670
- Brozek J., Grande F., Anderson J., Keys A. (1963): Densitometnic analysis of body composition: revision of some quantitative assumptions. Ann NY Acad Sci 1 10: 1 13
- Costill DL, Coyle EF, Fink WF, Lesmes GR, Witzmann FA (1979): Adaptations in skeletal muscle following strength training. J Appl Physiol 46:96
- Costill DL, Fink WJ, Pollock ML (1976): Muscle fiber composition and enzyme activities of elite distance runners. Med Sci Sports 8:96
- Dubowitz V. Brooke MH (1973): Muscle Biopsy: A Modern Approach. WB Saunders LTD. London, p 51
- Dudley GA, Staron RS, Murray TF, Hagerman FC, Luginbuhl A (1983): Muscle fiber composition and blood ammonia levels after intense exercise in humans. J Appl Physiol, 54:582
- Edgerton VR, Smith JL, Simpson DR (1975): Muscle fiber type populations of human leg muscles. Histochem J 7:259
- Eisenbeng BR, Kuda AM (1975): Stereological analysis of mammalian skeletal muscle. II. White vastus lateralis muscle of the adult guinea pig. J Ultrastruct Res 54:176
- Eisenberg BR, Kuda AM (1977): Retrieval of cryostat sections for comparison of histochemical and quantitative electron microscopy in a muscle fiber. J Histochem Cytochem 25:1169
- Essen B (1978): Glycogen depletion ofdiffenent fibre types in human skeletal muscle during intermittent and continuous exercise. Acta Physiol Scand 103:446
- Essen B, Hagenfeldt L, Kaijsen L (1977): Utilization of blood-borne and intramuscular substrates during continuous and intermittent exercise in man. J Physiol 265:489
- Essen B, Jansson E, Henriksson J, Taylor AW, Saltin B (1975): Metabolic characteristics of fibre types in human skeletal muscle. Acta Physiol Scand 95:153
- Evans WJ, Pinney SD, Young VR (1982): Suction applied to a muscle biopsy maximizes sample size. Med Sci Sports 14:101
- Gollnick PD (1982): Relationship of strength and endurance with muscle structure and metabolic potential. Int J Sports Med 3:26
- Goilnick PD, Armstrong RB, Saubert CW IV, Piehl K, Saltin B (1972): Enzyme activity and fiber composition in skeletal muscle of untrained and trained men. J Appl Physiol 33:3 12
- Green HJ (1978): Glycogen depletion patterns during continuous and intermittent ice skating. Med Sci Sports 10:183
- Green HJ, Daub B, Houston ME, Thomson JA, Fraser I, Ranney D (1981): Human vastus lateralis and gastrocnemius muscles. A comparative histochemical and biochemical analysis. J Neurol Sci 52:20 1
- Green HJ, Thomson JA, Daub WD, Houston ME, Ranney DA (1979): Fiber composition, fiber size and enzyme activities in vastus lateralis of elite athletes involved in high intensity exercise. Eur J Appl Physiol 41:109
- Haggmank T (1979): Hypotrophy of the soleus muscle in man after achilles tendon rupture. Discussion of findings obtained by computed tomography and morphologic studies. Am J Sports Med 7:12 1
- Hickson RC, Rosenkoetten MA, Brown MM (1980): Strength training effects on aerobic power and short-term endurance. Med Sci Sports 12:336
- Howald H (1982): Training-induced morphological and functional changes in skeletal muscle. Int J Sports Med 3:1
- Injger F ( 1977): A method for correlating ultrastructural and histochemical data from individual muscle fibers. Histochemistry 54:169
- Ingjer F (1979a): Effects of endunance training on muscle fiber ATPase activity, capillary supply and mitochondrial content in man. J Physiol 294:419
- Ingjen F (1979b): Capillary supply and mitochondnial content of different skeletal muscle fiber types in untrained and endurance-trained men. A histochemical and ultrastructural study. Eur J Appl Physiol 40: 197
- Jansson E, Kaijser L (1977): Muscle adaptation to extreme endurance training. Acta Physiol Scand 100:315
- Jansson E, Sjodin B, Tesch P ( 1978): Changes in muscle fiber type distribution in man after physical training. A sign of fiber transformation Acta Physiol Scand 104:235
- Johnson MA, Polgar J, Weightman D, Appleton D (1973): Data on the distribution of fiber types in thirty-six human muscles. An autopsy study. J Neurol Sci 18:111
- Kiessling K-H, Pilstrom L, Bylund A-Ch, Saltin B, Piehl K (1975):
- Morphometry and enzyme activities in skeletal muscle from middle-aged men after training and from alcoholics. In Metabolic Adaptation to Prolonged Physical Exercise. Edited by H Howald, JR Poortmans. Birkhausen Verlag, Basal, p 384
- Lexell J, Hanriksson-Larsen K, Sjostrom M ( 1983): Distribution of different fiber types in human skeletal muscles. 2. A study of cross-sections of whole m. vastus lateralis. Acta Physiol Scand 117 : 1 15
- Lithell H, OrlandenJ, Schele R, Sjodin B, Kanlsson J (1979): Changes in lipoportein-lipase activity and lipid stones in human skeletal muscle with prolonged heavy exercise. Acta Physiol Scand 107:257
- MacDougall JD, Sale DG, Moroz JR, Elder GCB, Sutton JR, Howald H (1979): Mitochondrial volume density in human skeletal muscle following heavy resistance training. Med Sci Sports 1 1:164
- Nygaard E, Sanchez J (1982): Intramuscular variation of fiber types in the brachial biceps and the lateral vastus muscles of elderly men: how representative is a small biopsy sample? Anat Rec 203:451
- Onlanden J, Kiessling K-H, KarlssonJ, Ekblom B (1977): Low intensity training, inactivity and resumed training in sedentary men. Acta Physiol Scand 101:35 1
- Prince FP, Hikida RS, Hagerman FC (1976): Human muscle fiber types in power lifters, distance runners and untrained subjects. Pfluegens Arch 363:19
- Prince FP, Hikida RS, Hagerman FC, Stanon RS, Allen WH (1981): A morphometric analysis of human muscle fibers with relation to fiber types and adaptations to exercise. J Neurol Sci 49:165
- Reichmann H, Pette D ( 1982): A comparative microphotometric study of succinate dehydnogenase activity levels in type I, IIA, and IIB fibers of mammalian and human muscles. Histochemistry 74:2 7
- Saltin B, Henniksson J, Nygaand E, Anderson P (1977): Fiber types and metabolic potentials of skeletal muscles in sedentary man and endurance runners. Ann NY Acad Sci 301:3
- Schantz P, Billeter R, Henriksson J, Jansson E (1982): Training induced increase in myofibrillar ATPase intermediate fibers in human skeletal muscle. Muscle Nerve 5:628
- Sjogaard G, Houston ME, Nygaand E, Saltin B (1978): Subgrouping
- Of fast-twitch fibres in skeletal muscles of man. Histochemistny 58:79
- Sjostrom M, Angquist K-A, Bylund A-C, Fniden J, Gustavsson L, Schersten T (1982b): Morphometric analyses of human muscle fiber types. Muscle Nerve 5:538
- Sjostrom M, Kidman 5, Henriksson-Larsen K, Angquist K-A (1982a): Z- and M-band appearance in different histochemically defined types of human skeletal muscle fibers. J Histochem Cytochem 30:1
- Stanon RS, Hikida RS, Hagerman FC (1983): Reevaluation of human skeletal muscle fast-twitch subtypes: evidence for a continuum. Histochemistry 78:33
- Thomson JA, Green HJ, Houston ME (1979): Muscle glycogen depletion patterns in fast-twitch fibre subgroups of man during submaximal and supramaximal exercise. Pfleugers Arch 379:105
- Thorstensson A, Hulten B, Dobeln W, Kanlsson J (1976): Effect of strength training on enzyme and fiber characteristics in human skeletal muscle. Acta Physiol Scand 96:392
- Weibel E, Kistler G, Scherle W (1966): Practical stereological methods for morphometric cytology. J Cell Biol 30:2 3
- Wilmone JH, Costill DL (1974): Semi-automated systems approach to the assessment of oxygen uptake during exercise. J Appl Physiol 36:6 18
- Wilmore JH, Vodak PA, Parr RB, Girandola RN, Billing JE (1980): Further simplification of a method for determination of residual lung volume. Med Sci Sports 12:216
Перевод А.А. Крестининой и А.В. Самсоновой
1983_Staron_перевод.pdf