Краткосрочная изокинетическая тренировка с высокой и низкой скоростью
Краткосрочная тренировка с высокой и низкой скоростью показала большую степень повреждения мышечных волокон при высокой скорости силовой тренировки. Скорость движений не влияла на гипертрофию медленных мышечных волокон I типа, однако влияла на гипертрофию мышечных волокон II типа. Большая гипертрофия наблюдалась в руке, выполняющей быстрые эксцентрические сокращения.

Shepstone, T. N. Short-term high- vs. low-velocity isokinetic lengthening training results in greater hypertrophy of the elbow flexors in young men / T.N. Shepstone, J.E. Tang, S. Dallaire, M.D. Schuenke, R. S. Staron, S.M. Phillips//J. Applied of Physiology, 2005.– V.98.–P. 1768-1776.
Шепстоун Т.Н.
КРАТКОСРОЧНАЯ ИЗОКИНЕТИЧЕСКАЯ ЭКСЦЕНТРИЧЕСКАЯ ТРЕНИРОВКА С ВЫСОКОЙ И НИЗКОЙ СКОРОСТЬЮ ПРИВОДИТ К БОЛЬШЕЙ ГИПЕРТРОФИИ МЫШЦ-СГИБАТЕЛЕЙ ПРЕДПЛЕЧЬЯ У МОЛОДЫХ МУЖЧИН
ABSTRACT
Мы выполнили два исследования, чтобы определить эффект программы силовых тренировок, состоявшей из быстрых и медленных эксцентрических сокращений на гипертрофию мышечных волокон. В первом исследовании, на молодых людях мы исследовали эффект быстрых (3,66 рад/с; Fast) или медленных (0,35 рад/с; Slow), изокнетических силовых эксцентрических упражнений на поперечное сечение мышечного волокна и всей мышцы (CSA) (двуглавая мышца плеча). Двенадцать исследуемых (23,8 ± 2,4 года; Mean±SD), выполняли максимальные силовые эксцентрические упражнения обеими руками в течение 8 недель (3 дня в неделю), в течение которого они тренировали одну руку в быстром растяжении, в то время как контралатеральная рука выполнила эквивалентное число сокращений с медленной скоростью. До (Pre) и после (Post) тренировки, была взята биопсия мышцы в центре брюшка двуглавой мышцы плеча и проанализированы для оценки типа волокна и площади поперечного сечения. Площадь поперечного сечения мышечных волокон I типа достоверно изменилась как в медленной, так и быстрой руке (P <0,05) Площадь поперечного сечения мышечных волокон IIa и IIx типов возросла в обеих руках, но большее возрастание было в руке, которая выполняла быстрые эксцентрические упражнения (P <0.05). Площадь поперечного сечения сгибателей предплечья возросла, как в медленной, так и быстрой руке, но в руке, выполняющей быстрые сокращения увеличение было большим (P = 0.06). Максимальная момент силы также увеличился в большей степени (P <0.05) в быстрой руке, независимо от скорости при которой осуществлялось тестирование.
В исследовании II, мы попытались дать объяснение большей гипертрофии, наблюдаемого в первом исследовании, исследуя как индикатор «размыв» Z-линии, который как мы предполагали, будет больше в Быстром сокращении. Девять мужчин (21,7 ± 2.4 года) выполнили «острую» работу (n = 30, 3 сета по 10 повторений/в сете) максимальных эксцентрических сокращений на Быстрых и Медленных скоростях, используемых в первом исследовании. Биопсии показали, что Быстрое эксцентрическое сокращение приводит к большему повреждению Z-полосы (185 ± 17 %; P <0.01), на квадратный мм поверхности мышцы, чем в Медленной руке. В заключение делаем вывод, что тренировка с использованием быстрого растяжения (3,66 рад/с) приводит к большей гипертрофии и приросту силы чем медленного (0,35 рад/с). Большая гипертрофия, наблюдаемая в Быстрой руке (I исследование), может быть связана с большим количеством поврежденного белка («размыв» Z-полосы; исследование II).
повреждение мышцы; время напряжения; разрушение Z-полосы
ВВЕДЕНИЕ
ГИПЕРТРОФИЯ СКЕЛЕТНОЙ МЫШЦЫ в результате силовых тренировок происходит из-за хронического суммирования периодов положительного баланса белка. Положительный баланс белка возникает из-за синергитического стимулирующего эффекта силовых тренировок и синтеза белка мышцы (6,7,48). Увеличение синтеза белка направлено на изменение и увеличение белка в клеточных структурах, в частности в миофибриллах и является критическим в процессе скелетной гипертрофии мышцы. Большинство форм силовых упражнений вызывает также разрушение ультраструктуры белков, обычно наблюдаемой как «размыв» Z-линии и нарушение расположения миофибрилл, которые больше связаны с удлинением, чем укорочением мышц ( 17,18).
И удлиняющие и укорачивающие сокращения [термины, используемые согласно аргументам, выделенным Faulkner (14), но часто называемые эксцентрическими и концентрические сокращениями, соответственно] вызывают то, что называется ультраструктурным повреждением белка, которое больше при эксцентрических сокращениях (17,18). При повторении эксцентрических сокращений, комбинация факторов вызывает уменьшение повреждения: так называемый эффект «повторения» (см. реф. 8) Исследования гипертрофии показали, что силовая тренировка мышцы, при использовании изотонического (21,26) и изокнетического (13,23,24,27,40) тренировочного протоколов в отсутствии удлинения мышцы приводит к меньшей гипертрофии, и меньшему приросту силы, чем соответствующая тренировка, состоящая исключительно из концентрических сокращений или комбинаций концентрического и эксцентрического сокращений. Однако, не всегда наблюдается большая гипертрофия при использовании изокинетической программы тренировок с эксцентрическими сокращениями (9,28,31). Было показано, что эксцентрические сокращения, выполняемые с большей скоростью приводят к большему увеличению силы, чем медленные (13,36), однако, механизм (ы), ответственный за это явление остается неизвестным. Используя ультразвук (B-способ), Farthing и Chilibeck (13) показали, что толщина мышцы (то есть гипертрофия), была больше при эксцентрических по сравнению с концентрическими сокращениями и что была тенденция (приблизительно на 5 % больше) для большей гипертрофии в быстрой (3,14 рад/с) руке по сравнению с медленной (0,52 рад/с). Используя такой смешанный проект, эти авторы не смогли обнаружить различия между быстрой и медленной рукой (13).
Мы выдвинули гипотезу, что тенденция к большей гипертрофии, замеченной при более быстрых эксцентрических упражнениях по сравнению с более медленными, связана с большим повреждением мышечных белков при быстрых сокращенииях (51,52).
Дополнительно, высокая скорость, достигаемая при удлинении мышцы, из-за механики мышцы на эксцентрической части кривой «скорость-сила», приводит к развитию большей силы мышц, которая также может стимулировать больший белковый синтез, приводящий к большей гипертрофии мышцы.
Цель настоящего исследования (исследование I) состояла в том, чтобы исследовать фазу ранней адаптации мышечного волокна и целой мышцы (CSA) сгибателей предплечья у молодых людей после изокнетический программы силовых тренировок с различными скоростями растяжения [низкая скорость (Медленная; 0,35 рад/с) и высокая скорость (Быстрая; 3,66 рад/с)]. Мы использовали нетренированных исследуемых, чтобы видеть максимальный ответ в терминах гипертрофии (2) Учитывая изменчивость в степени (32,49), так же как и во времени (20) гипертрофии, вызванной силовыми тренировками мы использовали краткосрочное исследование, и попытались обнаружить различия между двумя видами силовых тренировок. Мы хотели использовать программу, в которой сравниваются друг с другом разные конечности одного исследуемого, потому что такая программа лучше, чем при сравнении разных исследуемых и такая программа позволила бы увидеть эффект работы с различными скоростями Мы также решили проводить исследования на двуглавой мышце плеча, потому что исследуемые, при максимальном удлинении четырехглавой мышцы бедра смогли бы развить такие моменты, которые могли бы превысить максимальный вращающий момент, который могут оценить большинство динамометров (405 N·m в нашем случае), особенно после тренировок.
Мы выдвинули гипотезу, что тренировка с большой скоростью растяжения приведет к большему приросту силы (12,36) и гипертрофии (13). Большая гипертрофия в Быстрой руке должна была произойти, несмотря на то, что импульс силы под кривой «время–сила») имеет значительно меньшие значения, чем в медленной руке.
Чтобы исследовать потенциальные механизмы, лежащие в основе отличительных эффектов скорости на гипертрофию мышц, мы также провели отдельное исследование, в котором были исследованы изменения в разрушении Z-линии после эксцентрических сокращений с невысокой и высокой скоростью. Биопсии были взяты, у другой группы исследуемых, не принимавших участия в первом исследовании, чтобы исследовать степень разрушения Z-линии, вызванного единственным тренировочным занятием с использованием эксцентрических упражнений. Наша гипотеза состояла в том, что быстрое удлинение вызовет большее разрушение Z-линии, чем медленное удлинение, которое является индикатором разрушения белков и которое является предшественником гипертрофии мышцы (51,52).
МЕТОДЫ
Первое исследование. Тренировочная программа
Исследуемые. Двенадцать здоровых мужчин (возраст 23,8 ± 3,4 лет, рост 178,5 ± 9,6 см, вес 82,5 ± 10,3 кг), которые были двигательно активными (то есть, не тренировались) участвовали в 8-недельной тренировке. Исследуемые выполняли не более, чем 1-2 часовые тренировки в неделю структурированной физической деятельности, и ни один до этого не выполнял силовых упражнений. Исследуемые не принимали дополнительно никаких веществ, стимулирующих увеличение синтеза белка. Исследуемые прошли обычный медицинский осмотр, включающий анкетирование и оценку физического состояния и на этом основании считались здоровыми. Исследуемым сообщали о целях и рисках, связанных с исследованием, и они дали письменное согласие на основе полной информации. Проект был одобрен Правлением Этики McMaster Университета.
Общий экспериментальный протокол
За одну неделю до даты начала исследования, исследуемые ознакомились с тренажером (Biodex-Systems 3, Biodex Medical Systems, Shirly, NY). Перед началом тренировок у испытуемых определялась площадь поперечного сечения двуглавой мышцы плеча (в средней части) посредством компьютерного томографа (pQCT) высокой разрешающей способности. Исследуемые также подверглись ряду предварительных тестирований уровня силы (см. измерение силы) на обеих руках независимо друг от друга. Используя эту процедуру, и данные, согласно которым мышцы-сгибатели предплечья – легко возбудимые, поэтому легко достигают максимальной силы сокращения после небольшой тренировки (33), поэтому не удивительно, что мы наблюдали высокие межклассовые коэффициенты корреляции 0,95 и выше при повторных испытания максимального вращающего момента в пределах (r = 0,97) и между (r = 0,95) серией испытаний. Пять — семь дней спустя, у исследуемых были взяты образцы биопсии из средней части брюшка двуглавой мышцы плеча каждой руки. Выбор руки, выполняющей быстрые и медленные сокращения учитывал доминантный характер одной из рук.
Исследуемые начали тренировки после 1 недели отдыха. Для следующих 8 недель, исследуемые приходили в центр исследований каждый понедельник, среду, и пятницу для тренировок. В течение первой недели, выполнялся один сет (10 повторений в сете) максимальных эксцентрических сокращений для Быстрой и Медленной руки. Каждую последующую неделю добавлялся один сет с 180 с отдыха между сетами. К 4 неделе исследуемые выполняли четыре сета в каждый тренировочный день. В дальнейшем количество сетов не превышало четырех. Все исследуемые закончили 100 % предписанных 24 учебных сессий.
После 8 недель тренировок, исследуемым снова дали 4 дня отдыха перед последним тестированием, в котором оценивался максимальный момент силы. Биопсии мышцы были получены на расстоянии 4-5 см вперед или назад от первого места, где бралась биопсия.
Измерение силы. Измерение силы выполнялось до и после эксперимента в полностью случайном порядке с 4 мин отдыха между попытками на разных скоростях сокращения (0,35, 1,05, 2,10, 3,14, и 3,66 рад/с) и режимах сокращения (эксцентрический, концентрический, и изометрический). Исследуемые располагали локоть так, чтобы локтевой сустав соответствовал оси вращения динамометра Biodex, и они могли удобно схватить ручку рычага, в то время как их предплечье было в супинированном положении. Ограничившие ремни должны были обеспечить безопасность исследуемого при его работе на динамометре. Очень важно, ограничивающий ремень был помещен по диагонали к плечу, чтобы не допустить работу других мышц кроме мышц-сгибателей предплечья во время развития максимальных сокращений.
Изометрический момент силы. Исследуемые выполнили три повторения максимального произвольного сокращения при 1,05 рад (120 °) разгибания руки. Каждое сокращение было 5 с по продолжительности с 30 с отдыха между сокращениями. Максимальный изометрический момент силы, выбирался из этих трех сокращений
Концентрический момент силы. Концентрический момент силы был зарегистрирован как самый высокий пиковый вращающий момент трех повторений через 1,57 радиуса (90 °) диапазона движения. Каждое повторение началось с 3,05 рад (175 °) разгибания руки и заканчивалось, когда исследуемый сгибал руку, рычаг Biodex, с максимальной силой при угле 1,48 рад (85 °) сгибания локтя. Концентрический момент силы был оценен на пяти различных скоростях: 0,35, 1,05, 2,10, 3,14, и 3,66 рад/с.
Эксцентрический момент силы. Измерения эксцентрического момента силы были сделаны на пяти скоростях испытания как при измерении концентрического момента силы. Эксцентрический момент силы был взят от самого высокого пикового вращающего момента трех повторений, начинающихся при угле между плечом и предплечьем в 1,48 рад (85 °) и заканчивающихся в 3,05 рад (175 °). Каждое эксцентрическое сокращение, выполняемое на пяти скоростях (0,35, 1,05, 2,10, 3,14, и 3,66 рад/с), был закончено исследуемым, при максимальном сопротивлении рычага, который затем возвращался к исходному положению.
Компьютерная томография (pQCT). Средняя линия брюшка двуглавой мышцы плеча определялась как среднее расстояние между подмыщечной впадиной и antecubital. Рука была тогда вставлена в маленький цилиндр, который оценивает площадь измерения плотности pQCT (Stratec XCT 2000 Densiometer, Norland Medical System (Fort Atkinson, WI). Рука исследуемого оставалась неподвижной в течение приблизительно 4 минут, в то время как осуществлялось (2,5 мм) сканирование. Площадь поперечного сечения мышцы оценивалась посредством программного обеспечения фирмы BonAlyze (Jyvaskyla, Финляндия), которое автоматически идентифицирует ткань мышцы, оценивая ее плотность и геометрию. Программное обеспечение BonAlyze исключает межмышечный жир, определяя количество пикселов в пределах диапазона, соответствующего плотности мышцы (35-160 mg/cm3) и исключает пикселы, которые соответствуют плотности межмышечного жира. В нашей лаборатории, повторное сканирование середины двуглавой мышцы плеча одним и тем же исследователем давали относительную погрешность менее 1,33 % (12 повторных сканирований). При смене исследователя погрешность в оценке площади поперечного сечения мышцы не превышала 2,6 %. Следовательно, чтобы минимизировать изменчивость, все просмотры одного исследуемого были выполнены одним исследователем.
Биопсия мышцы. Образцы биопсии мышц были получены от каждого исследуемого под местной анестезией (2%-ый раствор лидокаина) при использовании ручного всасывания. Одна биопсия была взята в средней части правой и левой двуглавой мышцы плеча с целью установить базовый уровень. Второй образец биопсии был взят от каждой тренируемой руки в том же самом месте после эксперимента. Образцы немедленно анализировались, освобождались от видимого жира и соединительной ткани, и сохранялись при оптимальной температуре (OCT; Tissue Tech, Sakura Finetechnical) Разрез делался перпендикулярно ходу мышечных волокон. Образцы мышечных волокон были быстро заморожены в изопентане, охлажденном жидким азотом, и затем они были сохранены при температуре (-80°C) для последующего анализа.
Замороженные образцы мышечных волокон были последовательно поперечно разрезаны на криостате микротома толщиной в 10 мкм (модель HM500OM, MIKROM International, Waldorf, Германия) для гистохимического анализа. Гистохимический анализ на миофибриллярную АТФ-азу (mATPase) был выполнен, используя pH прединкубации равную 4,60 (50mM, ацетат калия и 17.5 mM хлорид кальция) в течение 6,5 минут, чтобы определить композицию мышечных волокон. Слайды были затем ополоснуты с дистиллированной водой и инкубировались в 3 mM ATФ при щелочном растворе (75 mM, глицин, 40,5 mM, хлорид кальция, 75 mM, NaCl, и 67,5 mM NaOH, приспособленный к pH равному 9,4) в течение 45 минут в 37°C и равномерно взбалтываемом в управляемом температурой шейкере инкубатора. После инкубации ATФ, было сделано полоскание дистиллированной водой, и образцы были выведены в 1%-ом CaCl2 в течение 3 минут комнатной температуры. Образцы были снова ополоснуты дистиллированной водой и инкубированы в 2%-ом СоCl2 в течение 3 минут при комнатной температуре. Затем следовало следующее полоскание с дистиллированной водой и инкубацией в 1%-ом сульфиде аммония в течение 1 минуты при комнатной температуре. Образцы были ополоснуты дистиллированной водой пять раз прежде, чем быть обезвоженным, находясь в течение 2 минут в концентрациях этанола (70, 80, 90, 95, и 100 %). Образцы были тогда очищены, используя ксилол. После того, как слайды были высушены, над ними были установлены сушилки, используя Permount (Fisher SP15) и очень быстро высушены.
Слайды рассматривались с использованием светового микроскопа (Olympic BX-60, Olympus America, Melvile, NY), после чего изображения были переведены в цифровую форму, используя SPOT-камеру (модель SP401-115, SPOT Diagnostic Instruments, Sterling Heigts, MI, и визуализируемое программное обеспечение SPOT (V3.2.4 для Windows, SPOT Diagnostic Instruments). Изображения были проанализированы при использовании обоих программных обеспечений Image-J (National Institut Mental Health, Bethesda, MD) и Imige Pro Plus (V4.0 для Windows, Media Cybernetics, Silver Spring, MD). Затем выполнялось 5-7 изображений, в 200 кратном усилении каждого образца, которое в значительной степени зависело от качества последовательных рассечений. Каждое изображение содержало приблизительно 30-50 волокон. Три волокна (I типа, IIa, и IIx) последовательно отличали, используя программное обеспечение Image-J, устанавливая пределы сокращения, приводящие к созданию оптической плотности «bins» согласно самому темному (I типа), самому светлому (тип IIa), и промежуточному (тип IIx) волокон, как предварительно описано (44). Классификация типа волокна таким образом зависит от интенсивности окрашивания в соответствии с mATPase гистохимическим протоколом. При pH равном 4,60, светлые, промежуточные и темные мышечные волокна соответствуют типу IIa волокна, типу IIx, и I типу, соответственно. Собранные изображения были преобразованы в 8-битовую шкалу, дающую 256 изображений шкалы яркости, которая линейно измеряет каждый пиксел и назначает ценность от 0 (черный) до 255 (белый). Устанавливая более низкие и высокие пороговые значения, оптические «bins» плотности были созданы: 0-95 для темных областей, 100-175 для промежуточных областей, 180-255 для светлых областей. Используя эти сокращения, были более объективно классифицированы три типа волокна. Подобная процедура использовалась в программном обеспечении Image Pro Plus. При сравнении способности программ Image-J и Image Pro Plus классифицировать одни и те же изображения (n= 26 образцов полученных до и после), мы получили r=0,985 (P<0,001), и мы также нашли, что коэффициент корреляции при повторном анализе изображений был равен 1,0 для обеих программ.
Прямые исследование на основе программного обеспечения Image Pro Plus позволили определить площадь поперечного сечения мышечных волокон, которая выражалась в квадратных мкм. Процент площади, занимаемый волокнами одного типа определялся как площадь поперечного сечения одного волокна определенного типа, разделенную на сумму площадей волокон всех типов и умноженную на 100. Распределение типа волокон (количество волокон одного типа как процент от общего количества исследованных волокон) было вычислено автоматически, используя программу Image Pro Plus.
Протокол MHC. анализ типов мышечных волокон на основе определения тяжелых цепей миозина (MHC) был выполнен как описано в (16). Кратко, четыре — шесть последовательных образцов мышцы разрезаны на слайды (20 мкм) и поместили в микрофуги трубы, содержащие 250 мл lising buffer (10 % wt/vol глицерин, 5 % vol/vol 2-mercaptoethanol, и 2.3 % wt/vol SDS в 62.5 mM pH 6.8), и были нагреты в течение 10 минут в 60°C. Приблизительно 4-6 мл разложенной таким образом мышцы были загружены в 20 см на 20 см на 1,5 мм поликриламидный гель. Гель размещался таким образом, что лучшие 25 % геля был 4 %, складывающие гель, тогда как остающиеся 75 % геля были градиентом акриламида на 4-8 %. Образцами управляли внезапно (19-21 часов) при 120 В и впоследствии окрашены синим цветом Coomassie. Три отдельных и отличных MHC изоформы (I, IIa, и IIx) были визуально идентифицированы согласно их массам (по сравнению с известными стандартами). Гели были тогда просмотрены, используя лазерный денситометр, относительная красящая интенсивность (то есть, число произвольных денситометрических единиц) каждой полосы была вычислена, и интенсивность была выражена как процент от полной красящей интенсивности (то есть, суммированные произвольные единицы всех трех полос).
Статистический анализ (исследование I). Размер мышечных волокон, тип волокна, pQCT, и данные MHC был проанализированы при использовании двух факторного анализа ANOVA, сопоставляя результаты, полученные до и после эксперимента, и условие работы (Быстрые и Медленные эксцентрические сокращения) как факторы. Данные силы были проанализированы при использовании четырех факторного анализа ANOVA, сопоставлялись результаты, полученные до и после, условие (быстрые и медленные эксцентрические сокращения, режим сокращение (эксцентрический и концентрический), и скорость (5 уровней: 0,35, 1,05, 2,10, 3,14, и 3,66 рад/с) как факторы. Все исследования были выполнены, используя SPSS (версия 11.5, Чикаго, IL). Статистическое значение для всех исследований было принято как P <0.05. Существенные главные эффекты и взаимодействия, где замечено в ANOVA, были далее проанализированы при использовании теста Тьюки. Представлены средние арифметические и стандартное отклонение (SD).
Исследование II: Острое Исследование
Исследуемые. Девять мужчин (возраст 23,2 ± 2,4 года, рост 181,9 ± 6,1 см, вес 81,1 ± 5,6 кг), которые были рекреационально активными (то есть, не выполняли никаких силовых тренировок и не более 1 тренировочного занятия в неделю) участвовали в остром протоколе исследования. Исследуемые должны были пройти обычный медицинский осмотр, основанный на анкетном опросе, и физической экспертизе и считались здоровыми. Исследуемым сообщали о целях и рисках, связанных с исследованием, и они дали письменное согласие на основе полной информации. Проект был одобрен Правлением Этики Университета McMaster.
Экспериментальный протокол. Перед исследованием в Центре исследуемых просили воздержаться от любой тяжелой деятельности, производимой верхними конечностями. В первый день эксперимента у исследуемых брали биопсию из брюшка двуглавой мышцы плеча правой и левой рук. Исследуемые также получили инструкцию относительно тренировки с использованием аппарата Biodex, который применялся для экспериментов. В течение недели после первой биопсии, исследуемых снова просили воздержаться от напряженной работы верхними конечностями. На следующий день, исследуемые в центре (Ivor Wynne Centre), должны были повторить протокол сокращения и впоследствии брался второй образец биопсии от каждой руки. Исследуемые снова возвратились, чтобы повторить протокол через 24 и 72 часа сокращения после второй сессии биопсии.
Экспериментальный протокол. Эксперимент проводился на динамометре Biodex. Исследуемые помещали локоть в Biodex, так, чтобы их предплечье было в супинированном положении, а локтевой сустав было на оси вращения рычага Biodex, и они могли удобно схватить ручку рычага. Ремень ограничения был помещен по диагонали, чтобы не допустить участие других групп мышц. Каждый исследуемый выполнил три сета (10 повторений/в сете) максимальных эксцентрических сокращений через 1,57 радиуса (90 °) окончания руки с 180 с отдыха между сетами. Одна рука выполняла движения с большой скоростью (3,66 рад/с), в то другой – медленно (0,35 рад/с).
Осуществление выборки мышцы. Образцы биопсии посредством иглы были получены от каждого исследуемый под местной анестезией (2%-ый лидокаин) при использовании ручного всасывания. Одна биопсия была взята от средней части правой и левой двуглавой мышцы плеча, чтобы установить базовый уровень. Другой образец биопсии был взят у каждого исследуемого таким же образом, как экспериментальный протокол с двумя сетами. Образцы немедленно освобождались от жира и соединительной ткани, и были размещены в охлажденный (4°C) фиксирующий (2 % gluteraldehyde buffered с натрием на 0.1 % cacodylate) для того, чтобы окрасить синим толуидином как описано предварительно (5,45,46).
Световая микроскопия. После начальной фиксации, образцы ткани были установлены в тетрооксиде осмия, обезвожены в градуируемых ваннах алкоголя этила, и вложенные в эпоксидную смолу (Spurr’s) с волокнами, ориентируемыми в длину. Каждый блок был тогда разделен (0,5 мкм) и окрашен посредством толуидина синим цветом.
Индивидуальные волокна от каждой продольной секции мышцы были изучены под 1000-кратным увеличением и исследованы на умеренное (3-10 непрерывных и/или смежных полос Z) и очень сильное (10 или больше непрерывных и/или смежных полос Z) повреждение Z-полос, как описано предварительно (5,45,46). Были вычислены средние площади, и количество «вытекания» Z-полосы было выражено на мм2 мышцы. Были вычислены площади образцов, при этом исследователь не знал, какие образцы он исследует (от быстрой или медленной мышцы). Коэффициент корреляции при повторном анализе 10 образцов был более 0,96. Кроме того, корреляция оценок «вытекания» Z-линии между исследователями используя этот метод также была больше 0,94, как сообщено предварительно (5,45,46). Чтобы уменьшить вариативность данных, один исследователь выполнил все исследования. Используя этот метод, показано, что секции, идентифицированные как наличие «вытекания» Z-полосы при использовании электронной микроскопии показывали также, что Z-полоса разрушена (5,45,46). Мы также подтверждали, используя электронную микроскопию (увеличение 3500-5000 раз), в 30 случайно отобранных блоках (6 блоков в покое, 12 блоков через 24 часа в быстрой руке, и 12 блоков через 24 часа в медленной руке), что «вытекание» Z-линии, было фактически областями ее разрушения. Повреждения, оцененные световым микроскопом и электронным микроскопом было сильно коррелированы (r= 0,98, P<0.0001).
Статистический анализ (II исследование). Вытекание Z-линии было проанализировано посредством парного t-теста, потому что в начале не было замечено ни одной разрушенной области, следовательно, полученные значения были по существу различием от базового уровня (то есть единственным различием между медленной и быстрой руками). Все исследования были выполнены, используя SPSS (версия 11.5). Статистическое значение для всех исследований было принято как P <0,05. Представленные значения – среднее и стандартное отклонение (SD).
РЕЗУЛЬТАТЫ
Первое исследование. Исследование посредством тренировочных занятий.
Сила. Тренировка привела к тому, что исследуемые были в состоянии произвести значительно большие максимальные моменты силы при эксцентрическом режиме сокращения мышцы, независимо от скорости, по сравнению с концентрическими сокращениями (P <0,05). Главный эффект тренировки состоял в том, что исследуемые были в состоянии произвести большие максимальные моменты силы после тренировок с высокой скоростью, независимо от скорости тестирования, по сравнению с концентрическими сокращениями (P = 0,023). Тренировка с высокой скоростью растяжения мышцы привела к увеличению максимума силы на 11,3 ± 10,4 N·m, тогда как тренировка с небольшой скоростью увеличила силу на 6,3 ± 12,0 N·m (P = 0.041; рис.1).

Изменение площади поперечного сечения всей мышцы до и после эксперимента показано на рис.2.

Значения площади поперечного сечения мышц до эксперимента значительно отличаются от нуля (P<0,01), существенного различия между руками (P= 0,060) нет, После эксперимента существуют существенные различия между руками (P= 0,024) работающими быстро и медленно, измеренные посредством post hoc тест.
Поперечное сечение мышечных волокон. Тренировка привела к существенному увеличению средней площади поперечного сечения всех типов волокон (P = 0,016). Мышечные волокна I типа, увеличились в размере (P = 0,042; рис.3) в среднем на 9,3 ± 5.0 % (вариация площади поперечного сечения волокон участвующих в медленных растяжениях от 3 до 560 мм2, быстрых – от 94 до 1300 мм2, без существенных различий между волокнами выполняющими быстрые растяжения и медленные растяжения (P= 0.19).
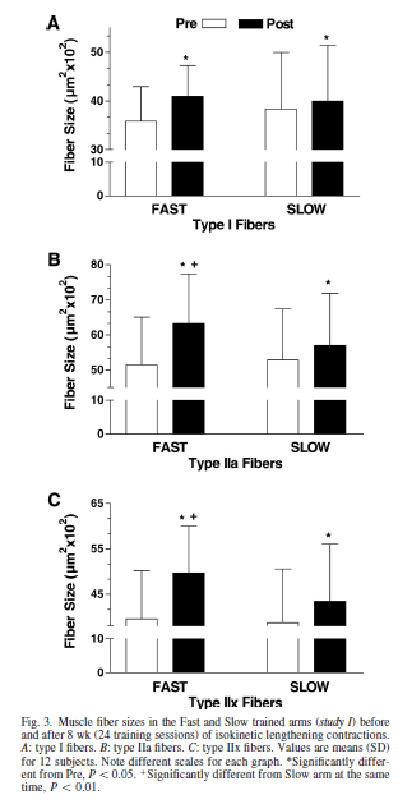
Изменение площади поперечного сечения мышечных волокон после тренировок только в волокнах II типа было больше в «Быстрой руке» (IIa: варьировало от = 279 до 1548 мм2; IIx варьировало от 963 до 1730 мм2). «Медленная рука» (IIa: варьировало от = 57-966 мм2; IIx варьировало от 205 до 1124 мм2; различия достоверны между всеми P= 0,007; рис.3
Тип мышечных волокон. Было замечено существенное уменьшение распределения волокон IIx типа (P = 0,031; таблица 1 независимо от тренировок. Хотя статистическое различие не было достигнуто, сохранялась тенденция к уменьшению занимаемой площади (в %) волокон типа IIx (P= 0,056; таблица 1).

Было, однако, существенное различие в площади (%), занимаемой волокнами IIa типа у руки, выполняющей быстрые эксцентрические сокращения до и после эксперимента (Р<0,05). Площадь, занимаемая волокнами IIa типа также была достоверно выше при выполнении движений быстрой рукой (P= 0.041).
Содержание MHC. Процент от изоформ MHC показал существенное уменьшение (в %) от типа IIx изоформ в быстрой и медленной руке после тренировок (P = 0,033; таблица 1). Точно так же было отмечено увеличение процента изоформ IIa типа после тренировок, которое было больше у быстрой руки по сравнению с медленной.

Процент изоформ MHC выраженный согласно электрофорезному гелю хорошо коррелировал с областью процента для каждого волокна, которые были проанализированы на основе mATPase гистохимического анализа (r= 0,95, P= 0.0006; рис.4).
Исследование II: Исследование «Вытекания Z-линии».
Степень умеренного разрушения Z-полосы (выраженная к мм2 мышцы), замеченная во всех образцах мышцы после тренировки была существенно больше (P <0.001) чем базовые образцы у которых не было никаких повреждений. Разрушение Z-полосы было больше (185 ± 17 %) после Быстрой тренировки по сравнению с медленной (P = 0,002; рис.5).

Степень чрезвычайных, по сравнению с умеренным, «вытеканий» Z-полосы (определенных как повреждение более 10 последовательно расположенных в длину или смежных саркомеров) (17,18) не сильно отличалась между медленной и быстрой рукой (P = 0,096) (Быстрая рука = 0,10 ± 0,15 и Медленный = 0.04 ± 0,11 области чрезвычайного повреждения к мм2 мышцы), но это было соблюдено в шести из девяти Быстрых образцов и только в двух из девяти Медленных образцов.
ОБСУЖДЕНИЕ
Результаты настоящего исследования показывают, что программа силовых тренировок, состоящая из быстрых изокнетических (3,66 рад/с) эксцентрических сокращений привела к большей гипертрофии мышечных волокон II типа, по сравнению с тренировками, выполненными с более медленной скоростью (0,35 рад/с). Это заключение основывается на значительно большем увеличением размера мышечных волокон, определенным по mATPase (миозин-АТФазе гистохимическим способом). Дополнительные данные о большей гипертрофии, наблюдаемой при выполнении быстрых эксцентрических тренировок демонстрировалась нашими данными, посредством измерениия площади поперечного сечения двуглавой мышцы плеча посредством компьютерной томографии. Хотя мы не наблюдали существенное различие между степенью изменения в Быстрой и Медленной руками посредством компьютерной томографии, имелась сильная тенденция к таким различиям (P = 0.06) между «Быстрой рукой», имеющей большую гипертрофию по сравнению с Медленной рукой. Вероятно из-за статистической ошибки второго рода, мы не сообщаем ни о каком различии в гипертрофии между Быстрой и Медленной руками, полученной посредством компьютерной томографии.
Множество исследований (13,21,23,24,26,27,49) показали, что тренировка мышцы, в которой использовались эксцентрические сокращения, приводит к большей гипертрофии по сравнению с концентрическими, или комбинации концентрических и эксцентрических сокращений. Мы также признаем, что существуют данные о том, что не всегда наблюдается большая гипертрофия при выполнении работы только в эксцентрическом режиме по сравнению с концентрическим или комбинированном концентрическим и эксцентрическим (9,28,31). Однако, мы полагаем, что существует кое-что, которое свойственно эксцентрическим сокращениям, что приводит к гипертрофии. Например, способность эксцентрических упражнений в вызове гипертрофии изящно продемонстрировано LaStayo с коллегами (30), которые использовали велоэргометрию для продуцирования гипертрофии, но это произошло только когда упражнение выполнено в эксцентрическом режиме. Эксцентрическая эргометрия вызвала 52%-ое увеличение площади мышечных волокон и 36%-ого увеличения изометрической силы ноги, тогда как традиционная концентрическая эргометрия не вызвала никакого изменения в размере волокна или силе. Кроме того, компенсационная гипертрофия после вызванной иммобилизацией атрофии также показала, что эксцентрические упражнения вызывают больший прирост силы и гипертрофии мышечных волокон (25). Недавно, Farthing и Chilibeck (13) нашли, что тренировка сгибателей предплечья с быстрыми эксцентрическими сокращениями (3,14 рад/с) в противоположность быстрому или медленному концентрическому сокращению (3,14 и 0,52рад/с) приводят к большей гипертрофии мышцы. Кроме того, различие в степени гипертрофии (5%), вызванной эксцентрическими упражнениями с быстрой и медленной (0,52 рад/с) скоростью было статистически недостоверно (13); их проект смешанного фактора вероятно испытывал недостаток в статистической мощности обнаружить это различие. В нашем исследовании, мы попытались уменьшить это влияние посредством модели двух рук у одного исследуемого и при использовании двух эксцентрических условий, которые были более несоизмеримы в скорости: 10-кратное превышение по сравнению с шестикратным, использованным Farthing и Chilibeck (13), думая, что большее различие в скорости может играть роль в выявлении различий в синтезе белка и как следствие – гипертрофии, согласно нашей гипотезе. Adams et al (1) недавно показал, что тренировка с использованием изометрических, концентрических и эксцентрических сокращений привела к эквивалентной гипертрофии у крыс. Интегралы момента силы для этих трех тренировочных способов были очевидно весьма отличны друг от друга. Несмотря на похожую степень гипертрофии, увеличения экспрессии mRNA выражения для фактора роста и инсулиноподобного фактора роста (IGF-I), два белка, которые были вовлечены как игроки в гипертрофический рост мышцы (19), были увеличены только в изометрических и концентрических условиях сокращения (1). У людей, единственное тренировочное занятие силовыми упражнениями в эксцентрическом режиме приводит к существенному возрастанию IGF-I и экспрессии mRNA по сравнению с тренировочным занятием, проводимом в концентрическом режиме (4). Мы наблюдали, несмотря на 10-кратный меньшее значение импульса силы (рис.6), большую степень гипертрофии, рис.3 с протоколом тренировки, в котором выполнялись эксцентрические сокращения с высоко скоростью.

Однако, почему мы наблюдали большую гипертрофию при выполнении быстрых эксцентрических упражнений по сравнению с медленными, хотя Adams с соавт (1), установил одинаковую гипертрофию при различных режимах сокращения и весьма несходными интегралами «сила-время» непонятно. Различие в модели, используемой в нашем исследовании в противоположность исследованию Adams с соавт (1) может объяснить некоторые из различий: крысы и люди; вызванные сокращения – произвольные сокращения; число тренировок было больше в нашем исследовании (24 тренировки у нас и только 10 тренировок у Adams с соавт. (1)).
Предварительно наша лаборатория маркировала ультраструктурное наблюдение смазанных или разрушенных Z линий как повреждение мышцы (5,46). Недавно было сообщено, что одно тренировочное занятие с эксцентрическими упражнениями приводящее к серьезному повреждению мышцы, не привело ни к каким обнаружимым беспорядкам миофибриллярной структуры белка (11). Фактически то, что мы наблюдали и маркировали как вытекание Z-линии, и предварительно назвали повреждением мышцы (5,46) на самом деле не было повреждением мышцы, а было «миофибриллярной реконструкцией». Это заключение было основано на изящном и детальном имммуногистологическом и электронно-микроскопическом исследовании состава белков аморфных или «мазавших» Z линии. Эти авторы показали, что «смазанные» Z линии поврежденной мышцы сильно отличались от нормы по составу белка. Они содержали выше нормы актин и десмин, что по заключению авторов было признаком ремоделирования белков мышечного волокна, а не повреждения (51,52). В этом случае наши данные указывают, на то, что при Быстрой тренировке то, которое, поскольку показало наше острое исследование, сопровождалось большим «вытеканием» Z-линии, рис. 5 будет связано с большим ремоделированием волокна. В нашей лаборатории недавно установлено, что эксцентрические сокращения, в противоположность концентрическим сокращениям, связаны с более быстрым повышением синтеза миофибриллярных белков (34). Если повышенный синтез белка является результатом повышенного accretion белка в течение долгого времени, согласно ряду событий, выделенных Phillips (37), то это обеспечивает основу того, почему мы видели большую гипертрофию при использовании тренировки с большой скоростью растяжения мышцы. Конечно, мы не имеем никаких прямых измерений обмена белка, синтеза или разрушения в этом исследовании, таким образом, эта теория остается гипотезой.
Тренировка увеличивала силу мышцы на всех тестируемых скоростях, независимо от способа тренировок; однако, большее значение максимального момента силы была больше в Быстрой руке во всех скоростях (то есть, главный эффект тренировки, рис.1). Этот результат наиболее вероятен по крайней мере частично последствие большей гипертрофии, замеченного в пределах Быстро-тренируемой руки (рис. 3).
Согласно предыдущим сообщениям (24,25,27) однако, прирост силы сильно зависит от способов тренировки. Следовательно, выполняя только эксцентрические сокращения, можно ожидать, что сила при эксцентрических сокращениях будет превышать силу при концентрических сокращениях. Есть также возможность переноса обучения, то есть, эффект одного протокола тренировки будет воздействовать на контралатеральную руку (10,41, 47). Однако, когда обе руки тренируются одновременно, невозможно оценить, как увеличение силы в одной конечности перешла бы к другой. В других исследованиях мы получили данные о том, что сила случайно варьировала в конечности, которая не тренировалась (41) или оставалась неизменной (47); следовательно, в нашем проекте, где обе конечности тренируются, кажется, что прирост силы, которая переходит от одной конечности к другой был бы минимальным, по сравнению с передачей силы к нетренируемой конечности. Мы признаем, что способ тренировки, возможно, был передан контралатеральной конечности, но свидетельство для этого явления в одновременном обучении конечностей указывает на одинаковые результаты к тому, что мы наблюдали, это — полное общее увеличение силы при удлинении мышцы без эффекта специфики для скорости Дополнительно, потому что прирост максимального момента силы был больше в Быстро-тренируемой руке, тогда любой перенос в терминах прироста силы должен был бы увеличить прирост силы в Медленно-тренируемой руке, в которой мы наблюдали «низший» прирост силы (рис. 1). Следовательно, тот факт, что мы видели различие между Быстрой и Медленной рукой в терминах прироста силы, еще более внушителен и вероятен из-за нашего использования одно исследуемого.
Полный прирост момента силы был весьма умеренным, и следовательно различия будет более трудно обнаружить и представить. Мы подозреваем, что умеренный прирост силы, возможно, произошел из-за некоторой остаточной усталости непосредственно от тренировок, несмотря на несколько дней отдыха после последней тренировки исследуемых перед оценкой силы. Фактически, рис. 7 показывает средний момент силы по неделе-к-недельному курсу исследования и показывает, что, в течение тренировок, еженедельный пиковый момент силы был фактически понижен, вероятно в результате ремоделирования белка мышцы (см. реферат 50), в течение некоторого времени перед восстановлением и в конечном итоге превысил до тренировочные значения только на последней неделе.

Этот пример пониженной силы и все же существенной модернизации волокна, в конечном счете приводящей к гипертрофии, подобен хроническому возбуждению низкой частотой у животных. В этой модели чрезвычайной перегрузки, постоянно понижается сила, и все же адаптация волокна все еще происходит, в то время как присутствует существенное ремоделирование волокна (22). В двух других исследованиях, в которых отличались скорости в эксцентрическом режиме, используемые для тренировки (12,36) прирост максимального момента силы был небольшим. Например, Paddon-Jones с соавт (36) не нашел никакого существенного прироста максимального момента силы в медленно-тренируемом удлиняющем сокращении (0,52 рад/с) и увеличение на 5-10 N·m в быстротренируемомй руке (3,14 рад/с). Farthing и Chilibeck также сообщили, что приращения в максимальном моменте были в диапазоне 11-14 N·m.
Изменение типа мышечных волокон, которые обычно происходят при силовых тренировках, обычно демонстрировали изменение от IIx до волокон IIa (25,42,43). Однако, недавнее наблюдение распределения типа волокна после тренировок в эксцентрическом режиме с большой скоростью показало увеличение процента волокон IIx типа и уменьшения волокон I типа (36). При завершении существующего исследования, таких изменений найдено не было. Фактически, уменьшение было замечено в волокнах типа IIx и после Быстрой и Медленной эксцентрической тренировки и согласно mATPase гистохимического анализа и согласно гелю MHC электрофоретического анализа. Хотя предполагаемое сопутствующее увеличение % волокон IIa типа не было замечено, мы действительно наблюдали существенное увеличение общей площади, занятой волокнами IIa типа, но только в Быстро-тренируемой руке. Изменение от IIx до волокон IIa должно было ожидаться согласно Andersen и др. (3), которые выдвинули гипотезу, что тип IIx изоформ MHC «по умолчанию», часто выражен у нетренированных исследуемых. Однако, после силовых тренировок, MHC изоформ изменяются и увеличивается процентное содержание волокна типа IIa (25,42,43). Волокна IIa типа, хотя имеют более низкий максимальный момент силы (29), имеют большую окислительную способность, которая позволяет им противостоять утомлению. Кроме того, увеличения площади поперечного сечения мышечных волокон IIa и IIx типа дает значительно большую компенсацию, чем любое маленькое изменение от IIx до IIa.
Чтобы сопоставить наши данные с другими протоколами силовой тренировки, интересно отметить, что степень гипертрофии, которую мы наблюдали по сравнению с другими исследованиями, в которых тренировались сгибатели предплечья, показали подобные увеличения площади поперечного сечения мышцы (13), большие увеличения площади поперечного сечения и индивидуальной площади поперечного сечения мышечных волокон область мышечных волокон с более длительной тренировкой (35,38) или никаких изменений в размере мышечных волокон у тренируемых исследуемых, которые добавляли дополнительные сгибания на вершине своей тренировки. (2). Увеличение площади поперечного сечения мышцы (CSA) и площади мышечных волокон, которую наблюдали в нашей лаборатории и в других исследовательских центрах (13,35,38) имеют тенденцию быть большими, чем замеченное в таких мышцах как vastus lateralis (42,43). Следовательно, полученные нами данные должны рассматриваться в контексте краткосрочного протокола тренировок сгибателей предплечья и могли отличаться, если исследуемые были тренированными или изучались другие мышцы (то есть, мышцы нижних конечностей).
Мы заметили, что более высокая скорость (3,66 рад/с) изокинетических эксцентрических сокращений связана с большей мышечной гипертрофией, чем низкая (0,35 рад/с) скорость эксцентрических упражнений. Это произошло несмотря на более чем 10-кратное большее время под напряжением в медленной по сравнению с быстрой рукой. Мы заметили, что даже один сет быстрых эксцентрических сокращений привел к большему разрушению ультраструктуры мышцы, которая свидетельствовала о большем ремоделировании белка с потенциально более быстрым началом (34) и потенциально большей продолжительностью синтеза белка. В течение долгого времени, больший синтез белка может привести к большей гипертрофии.
СНОСКИ
Address for reprint requests and other correspondence: S. M. Phillips, Dept. of Kinesiology, IWC AB116, McMaster Univ., 1280 Main St. W., Hamilton, ON, Canada L8S 4K1 (E-mail: phillis@mcmaster.ca)
ЛИТЕРАТУРА
- Adams GR, Cheng DC, Haddad F, and Baldwin KM. Skeletal muscle hypertrophy in response to isometric, lengthening, and shortening training bouts of equivalent duration. J Appl Physiol 96: 1613–1618, 2004.
- Alway SE, Grumbt WH, Stray-Gundersen J, and Gonyea WJ. Effects of resistance training on elbow flexors of highly competitive bodybuilders. J Appl Physiol 72: 1512–1521, 1992.
- Andersen JL, Mohr T, Biering-Sorensen F, Galbo H, and Kjaer M. Myosin heavy chain isoform transformation in single fibres from m. vastus lateralis in spinal cord injured individuals: effects of long-term functional electrical stimulation (FES). Pflügers Arch 431: 513–518, 1996.
- Bamman MM, Shipp JR, Jiang J, Gower BA, Hunter GR, Goodman A, McLafferty CL Jr, and Urban RJ. Mechanical load increases muscle IGF-I and androgen receptor mRNA concentrations in humans. Am J Physiol Endocrinol Metab 280: E383–E390, 2001.
- Beaton LJ, Tarnopolsky MA, and Phillips SM. Contraction-induced muscle damage in humans following calcium channel blocker administration. J Physiol 544: 849–859, 2002.
- Biolo G, Tipton KD, Klein S, and Wolfe RR. An abundant supply of amino acids enhances the metabolic effect of exercise on muscle protein. Am J Physiol Endocrinol Metab 273: E122–E129, 1997.
- Borsheim E, Tipton KD, Wolf SE, and Wolfe RR. Essential amino acids and muscle protein recovery from resistance exercise. Am J Physiol Endocrinol Metab 283: E648–E657, 2002.
- Clarkson PM and Hubal MJ. Exercise-induced muscle damage in humans. Am J Phys Med Rehabil 81: S52–S69, 2002.
- Colliander EB and Tesch PA. Effects of eccentric and concentric muscle actions in resistance training. Acta Physiol Scand 140: 31–39, 1990.
- Colliander EB and Tesch PA. Effects of detraining following short term resistance training on eccentric and concentric muscle strength. Acta Physiol Scand 144: 23–29, 1992.
- Crameri RM, Langberg H, Magnusson P, Jensen CH, Schroder HD, Olesen JL, Suetta C, Teisner B, and Kjaer M. Changes in satellite cells in human skeletal muscle after a single bout of high intensity exercise. J Physiol 558: 333–340, 2004.
- Farthing JP and Chilibeck PD. The effect of eccentric training at different velocities on cross-education. Eur J Appl Physiol 89: 570–577, 2003.
- Farthing JP and Chilibeck PD. The effects of eccentric and concentric training at different velocities on muscle hypertrophy. Eur J Appl Physiol 89: 578–586, 2003.
- Faulkner JA. Terminology for contractions of muscles during shortening, while isometric, and during lengthening. J Appl Physiol 95: 455–459, 2003.
- Friden J, Sjöstrom M, and Ekblom B. Myofibrillar damage following intense eccentric exercise in man. Int J Sports Med 4: 170–176, 1983.
- Fry AC, Allemeier CA, and Staron RS. Correlation between percentage fiber type area and myosin heavy chain content in human skeletal muscle. Eur J Appl Physiol 68: 246–251, 1994.
- Gibala MJ, Interisano SA, Tarnopolsky MA, Roy BD, MacDonald JR, Yarasheski KE, and MacDougall JD. Myofibrillar disruption following acute concentric and eccentric resistance exercise in strength-trained men. Can J Physiol Pharmacol 78: 656–661, 2000.
- Gibala MJ, MacDougall JD, Tarnopolsky MA, Stauber WT, and Elorriaga A. Changes in human skeletal muscle ultrastructure and force production after acute resistance exercise. J Appl Physiol 78: 702–708, 1995.
- Goldspink G. Changes in muscle mass and phenotype and the expression of autocrine and systemic growth factors by muscle in response to stretch and overload. J Anat 194: 323–334, 1999.
- Green H, Goreham C, Ouyang J, Ball-Burnett M, and Ranney D. Regulation of fiber size, oxidative potential, and capillarization in human muscle by resistance exercise. Am J Physiol Regul Integr Comp Physiol 276: R591–R596, 1999.
- Hather BM, Tesch PA, Buchanan P, and Dudley GA. Influence of eccentric actions on skeletal muscle adaptations to resistance training. Acta Physiol Scand 143: 177–185, 1991.
- Hicks A, Ohlendieck K, Gopel SO, and Pette D. Early functional and biochemical adaptations to low-frequency stimulation of rabbit fast-twitch muscle. Am J Physiol Cell Physiol 273: C297–C305, 1997.
- Higbie EJ, Cureton KJ, Warren GL, and Prior BM. Effects of concentric and eccentric training on muscle strength, cross-sectional area, and neural activation. J Appl Physiol 81: 2173–2181, 1996.
- Hortobagyi T, Barrier J, Beard D, Braspennincx J, Koens P, Devita P, Dempsey L, and Lambert J. Greater initial adaptations to submaximal muscle lengthening than maximal shortening. J Appl Physiol 81: 1677–1682, 1996.
- Hortobagyi T, Dempsey L, Fraser D, Zheng D, Hamilton G, Lambert J, and Dohm L. Changes in muscle strength, muscle fibre size and myofibrillar gene expression after immobilization and retraining in humans. J Physiol 524: 293–304, 2000.
- Hortobagyi T, Devita P, Money J, and Barrier J. Effects of standard and eccentric overload strength training in young women. Med Sci Sports Exerc 33: 1206–1212, 2001.
- Hortobagyi T, Hill JP, Houmard JA, Fraser DD, Lambert NJ, and Israel RG. Adaptive responses to muscle lengthening and shortening in humans. J Appl Physiol 80: 765–772, 1996.
- Jones DA and Rutherford OM. Human muscle strength training: the effects of three different regimens and the nature of the resultant changes. J Physiol 391: 1–11, 1987.
- Larsson L, Edstrom L, Lindegren B, Gorza L, and Schiaffino S. MHC composition and enzyme-histochemical and physiological properties of a novel fast-twitch motor unit type. Am J Physiol Cell Physiol 261: C93–C101, 1991.
- Lastayo PC, Pierotti DJ, Pifer J, Hoppeler H, and Lindstedt SL. Eccentric ergometry: increases in locomotor muscle size and strength at low training intensities. Am J Physiol Regul Integr Comp Physiol 278: R1282–R1288, 2000.
- Mayhew TP, Rothstein JM, Finucane SD, and Lamb RL. Muscular adaptation to concentric and eccentric exercise at equal power levels. Med Sci Sports Exerc 27: 868–873, 1995.
- McCall GE, Byrnes WC, Dickinson A, Pattany PM, and Fleck SJ. Muscle fiber hypertrophy, hyperplasia, and capillary density in college men after resistance training. J Appl Physiol 81: 2004–2012, 1996.
- Moore DR, Burgomaster KA, Schofield LM, Gibala MJ, Sale DG, and Phillips SM. Neuromuscular adaptations in human muscle following low intensity resistance training with vascular occlusion. Eur J Appl Physiol 92: 399–406, 2004.
- Moore DR, Phillips SM, Babraj JA, Smith K, and Rennie MJ. Myofibrillar and collagen protein synthesis in human skeletal muscle after maximal shortening and lengthening contractions. Am J Physiol Endocrinol Metab. In press.
- O’Hagan FT, Sale DG, MacDougall JD, and Garner SH. Response to resistance training in young women and men. Int J Sports Med 16: 314–321, 1995.
- Paddon-Jones D, Leveritt M, Lonergan A, and Abernethy P. Adaptation to chronic eccentric exercise in humans: the influence of contraction velocity. Eur J Appl Physiol 85: 466–471, 2001.
- Phillips SM. Protein requirements and supplementation in strength sports. Nutrition 20: 689–695, 2004.
- Roman WJ, Fleckenstein J, Stray-Gundersen J, Alway SE, Peshock R, and Gonyea WJ. Adaptations in the elbow flexors of elderly males after heavy-resistance training. J Appl Physiol 74: 750–754, 1993.
- Roth SM, Martel GF, Ivey FM, Lemmer JT, Tracy BL, Hurlbut DE, Metter EJ, Hurley BF, and Rogers, MA. Ultrastructural muscle damage in young vs. older men after high-volume, heavy-resistance strength training. J Appl Physiol 86: 1833–1840, 1999.
- Seger JY, Arvidsson B, and Thorstensson A. Specific effects of eccentric and concentric training on muscle strength and morphology in humans. Eur J Appl Physiol Occup Physiol 79: 49–57, 1998.
- Shima N, Ishida K, Katayama K, Morotome Y, Sato Y, and Miyamura M. Cross education of muscular strength during unilateral resistance training and detraining. Eur J Appl Physiol 86: 287–294, 2002.
- Staron RS, Karapondo DL, Kraemer WJ, Fry AC, Gordon SE, Falkel JE, Hagerman FC, and Hikida RS. Skeletal muscle adaptations during early phase of heavy-resistance training in men and women. J Appl Physiol 76: 1247–1255, 1994.
- Staron RS, Leonardi MJ, Karapondo DL, Malicky ES, Falkel JE, Hagerman FC, and Hikida RS. Strength and skeletal muscle adaptations in heavy-resistance-trained women after detraining and retraining. J Appl Physiol 70: 631–640, 1991.
- Stewart BG, Tarnopolsky MA, Hicks AL, McCartney N, Mahoney DJ, Staron RS, and Phillips SM. Treadmill training-induced adaptations in muscle phenotype in persons with incomplete spinal cord injury. Muscle Nerve 30: 61–68, 2003.
- Stupka N, Lowther S, Chorneyko K, Bourgeois JM, Hogben C, and Tarnopolsky MA. Gender differences in muscle inflammation after eccentric exercise. J Appl Physiol 89: 2325–2332, 2000.
- Stupka N, Tarnopolsky MA, Yardley NJ, and Phillips SM. Cellular adaptation to repeated eccentric exercise-induced muscle damage. J Appl Physiol 91: 1669–1678, 2001.
- Tesch PA, Ekberg A, Lindquist DM, and Trieschmann JT. Muscle hypertrophy following 5-week resistance training using a non-gravity-dependent exercise system. Acta Physiol Scand 180: 89–98, 2004.
- Tipton KD, Borsheim E, Wolf SE, Sanford AP, and Wolfe RR. Acute response of net muscle protein balance reflects 24-h balance after exercise and amino acid ingestion. Am J Physiol Endocrinol Metab 284: E76–E89, 2003.
- Wang N, Hikida RS, Staron RS, and Simoneau JA. Muscle fiber types of women after resistance training—quantitative ultrastructure and enzyme activity. Eur J Appl Physiol 424: 494–502, 1993.
- Warren GL, Ingalls CP, Lowe DA, and Armstrong RB. Excitation-contraction uncoupling: major role in contraction-induced muscle injury. Exerc Sport Sci Rev 29: 82–87, 2001.
- Yu JG, Carlsson L, and Thornell LE. Evidence for myofibril remodeling as opposed to myofibril damage in human muscles with DOMS: an ultrastructural and immunoelectron microscopic study. Histochem Cell Biol 121: 219–227, 2004.
- Yu JG and Thornell LE. Desmin and actin alterations in human muscles affected by delayed onset muscle soreness: a high resolution immunocytochemical study. Histochem Cell Biol 118: 171–179, 2002.
Эта статья процитирована в следующих статьях:
- J. Tannerstedt, W. Apro, and E. Blomstrand Maximal lengthening contractions induce different signaling responses in the type I and type II fibers of human skeletal muscle J Appl Physiol, April 1, 2009; 106(4): 1412 — 1418.
- L. Holm, S. Reitelseder, T. G. Pedersen, S. Doessing, S. G. Petersen, A. Flyvbjerg, J. L. Andersen, P. Aagaard, and M. Kjaer Changes in muscle size and MHC composition in response to resistance exercise with heavy and light loading intensity J Appl Physiol, November 1, 2008; 105(5): 1454 — 1461.
- J. E. Tang, J. G. Perco, D. R. Moore, S. B. Wilkinson, and S. M. Phillips Resistance training alters the response of fed state mixed muscle protein synthesis in young men Am J Physiol Regulatory Integrative Comp Physiol, January 1, 2008; 294(1): R172 — R178.
- J. W Hartman, J. E Tang, S. B Wilkinson, M. A Tarnopolsky, R. L Lawrence, A. V Fullerton, and S. M Phillips Consumption of fat-free fluid milk after resistance exercise promotes greater lean mass accretion than does consumption of soy or carbohydrate in young, novice, male weightlifters Am. J. Clinical Nutrition, August 1, 2007; 86(2): 373 — 381.
- D. R. Moore, N. C. Del Bel, K. I. Nizi, J. W. Hartman, J. E. Tang, D. Armstrong, and S. M. Phillips Resistance Training Reduces Fasted- and Fed-State Leucine Turnover and Increases Dietary Nitrogen Retention in Previously Untrained Young Men J. Nutr., April 1, 2007; 137(4): 985 – 991
- T. C. Chen, K. Nosaka, and P. Sacco Intensity of eccentric exercise, shift of optimum angle, and the magnitude of repeated-bout effect J Appl Physiol, March 1, 2007; 102(3): 992 – 999
С уважением, А.В. Самсонова