Активность ферментов и состав мышечных волокон в скелетных мышцах
Активность ферментов и состав мышечных волокон изучались посредством биопсии. Исследовались: латеральная широкая мышца бедра и дельтовидная мышца. В исследовании приняли участие 74 нетренированных и тренированных испытуемых. МВ ST типа преобладали у спортсменов тренирующихся на выносливость. Приведены данные о проценте МВ ST типа и значениях МПК у спортсменов высокой квалификации.

Gollnick, P.D. Enzyme activity and fiber composition in skeletal muscle of untrained and trained men / P.D. Gollnick, R.B. Armstrong, C.W. Saubert IV, K. Piehl, and B. Saltin // Journal of Applied Phisiology, 1972.- Vol. 33.- No 3.- P. 312- 319.
Голльник П.Д.
АКТИВНОСТЬ ФЕРМЕНТОВ И СОСТАВ ВОЛОКНА В СКЕЛЕТНЫХ МЫШЦАХ НЕТРЕНИРОВАННЫХ И ТРЕНИРОВАННЫХ ЛЮДЕЙ
Посредством биопсии образцов из латеральной широкой и дельтовидной мышцы 74 нетренированных и тренированных людей изучалась активность сукцинатдегидрогеназы (СДГ) и фосфофруктокиназы (ФФК) и осуществлялась гистохимическая идентификация типов волокон и локализация окислительной деятельности. Активность СДГ была наивысшей в мышцах людей, участвующих в тренировке на выносливость. Наивысшая активность проявилась в мышцах, которые были широко вовлечены в работу на выносливость. Только незначительные различия были замечены в деятельности ФФК. Только медленно сокращающиеся (МВ ST типа) и быстро сокращающиеся (МВ FT типа) волокна были идентифицированы в образцах мышц. МВ ST типа преобладали в мышцах спортсменов на выносливость. Во всех группах наблюдалась широкая вариация волокон. Установлено, что распределение волокна указывает на долю площади, занимаемой волокном в мышцах. Окислительная способность обоих типов волокон была больше у выносливых спортсменов, чем у других. Мышечный гликоген является наиболее высоким у тренированных людей. Нет закономерности для накопления гликогена для двух типов волокон.
Ключевые слова: сукцинатдегидрогеназа; гликоген; типы волокон; фосфофруктокиназа; скелетные мышцы человека
По меньшей мере, три различных типа волокон определены в большинстве видов скелетных мышц. Несколько систем было предложено для идентификации этих видов волокон (3, 5, 10, 24). В мышцах человека большинство исследователей выявили только два отчетливо различающихся типа волокна. Они были классифицированы как тип I и тип II (7, 16) или красные и белые (9) волокна. Edström и Nyström (9) отметили различия в процентном распределении этих двух видов волокон между различными мышцами и в одних и тех же мышцах у разных субъектов.
Взаимопревращаемость типов волокон была продемонстрирована у животных после перекрестной иннервации (8). Barnard и соавторы (3) также сообщили, что физическая тренировка привела к увеличению процента красных волокон и сокращению белых волокон в мышцах морской свинки. У животных, тренировки на выносливость могут изменить метаболические характеристики скелетных мышц, повышая активность некоторых ферментов цикла Кребса, митохондриальную концентрацию белка, и способность к окислению жира (11, 12, 14). Некоторые из этих изменений были отмечены в человеческих скелетных мышцах после тренировки (15, 26). Тем не менее, метаболические характеристики и состав волокна в мышцах тренируемых и нетренируемых людей и их отношение к деятельности определенных групп мышц, к их силовой характеристике не были изучены. Целью этого исследования являлось приближение к этой проблеме, изучая группы испытуемых, которые занимались различными видами тренировок, с разной интенсивностью тренировок в течение длительного периода времени.
ОБЪЕКТ И МЕТОДЫ ИССЛЕДОВАНИЯ
В исследовании участвовали девять групп мужчин (таблица 1). Были выбраны представители разных возрастных групп и состояний физической подготовки, принимавшие участие в различных спортивных мероприятиях, которые использовали различные группы мышц и учебные программы. В общей сложности были изучены характеристики 74 мужчин в возрасте от 17 и 58 лет.
Образцы латеральной широкой и дельтовидной мышцы были получены техникой пункционной биопсии (4) и были разделены на три части. Одна часть образца сразу же была заморожена в жидком азоте и хранилась в сухом льду для последующего гистохимического анализа. Остальные две части были взвешены и использованы для определения активности янтарной кислоты сукцинатдегидрогеназы (SDH) и фосфофруктокиназы (PFK) в 25 ° С методами Cooperstein с соавт. (6) и Shonk и Boxer (23), соответственно. В некоторых случаях биопсия была проведена повторно, чтобы обеспечить достаточное количество ткани для различных измерений. В таких случаях биопсия была сделана в том же самом месте отбора проб, как и первый раз.
Образцы, замороженные для гистохимического анализа, были исследованы под препаровочной лупой в холодном помещении при -25 C, для определения ориентации волокон. Затем они были охлаждены до температуры жидкого азота, помещены на держатели образцов в ОСТ заливочную среду (Ames Tissue-Tek) в температуру около 0 ° C, и сразу же погружены в жидкий азот. Последовательные сечения, толщиной 10 мк, были разрезаны в криостате при — 20° С и помещены на стекла для окрашивания. Активность аденозинтрифосфатазы (АТФаза) миозина, дифосфопиридин нуклеотид-диафоразы (ДПН-Н-диафораза) и активность альфа-глицерофосфатдегидразы оценивались методами Padykula и Herman (20), Novikoff с соавт. (18) и Wattenberg и Leong (27), соответственно. Распределение гликогена (толщиной 16 мк) в одном из последовательных сечений оценивалось реакцией Шифф-йодной кислотою (PAS) (21).
Фотографии образцов были сделаны 20 х 25 см черно-белыми отпечатками, с целью классификации волокна и оценки распределения окислительных и гликолитических волокон.
Количество гликогена в некоторых образцах определяли способом, описанным Karlsson и соавт. (13). Рибонуклеиновая кислота (РНК) была определена в образцах некоторых мышечных групп модифицированным методом Schmidt-Thannhauser, описанным Munro и Fleck (17). Дрожжевая РНК была использована в качестве стандарта.
ТАБЛИЦА 1 Скелетные мышцы тренируемых и нетренируемых людей. Физические характеристики, число волокон, активность ферментов и МПК в исследуемых группах
| Группа, возраст, номер | Рост, вес | Тип работы (Рука-нога) | % содержание МВ ST типа | Активность ферментов, µмоль / г мин -¹ | МПК мл/ кгмин-¹ | |
| SDH | PFK | |||||
| Нетренируемые, 27 (24-30) лет, n = 12 | 179 см (166-187) 76.5 кг (66.4-105.5)
| Р Н | 46.0 ± 6.8 (14.3-59.8) 36.1±5.0 (13 .0-50.8)
| 3.6 ± 0.4 (2.1-6.0) 4.3 ± 0.6 (2.7-6.5) | 21.6 ± 3.7 (19.8-28 .6) 25.3 ± 2.1 (17.7-38.0)
|
42.9 ± 1.9 (32-53) |
| Нетренируемые, 39 (31-52) лет, n = 14 | 178 см (168-191) 75.6 кг (63.2-88.6) | Р Н
| 45.2 ± 2.7 (33.5-58.3) 43.9 ± 4.8 (24.0-72.9) | 3.5±0.4 (1.9-6.6) 4.4 ± 0.3 (2.3-6.5) | 23.0 ± 1.9 (16.5-32.3) 25.2 ± 1 .9 (14.3-36.6) |
41.4 ± 1.2 (35-49) |
| Тренеруемые,* 25 (17-30) лет, n = 12 | 179 см (165-189) 68.8 кг (59.6-76.0) | Р Н | 54.8 ±3.4 (45.5-66.2) 52.3 ± 6.8 (26.0-62.5) | 4.0 ± 0.5 (2.7-5.3) 6.0 ±0.3 (4 .0-8 .0) | 19.1 ± 1.8 (13.3-24.6) 19.9 ± 3.6 (14.3-28.3) |
57.1 ± 1.3 (51-65) |
| Велосипедисты, 24 (18-33) лет, n = 4 | 182 см (175-189) 74.5 кг (74 .0-75 .0) | Р Н
| 50.7 ± 4.4 (39.5-63.8) 61.4 ± 5.9 (47.9-73 .2) | 6.1 ± 0.2 (5.5-6.7) 11.0±1.0 (8.2-12.4) | 24.0 ± 4.0 (19.4-32 .0) 23.9 ±1.4 (21 .4-32 .0) |
68.2 ± 2.2 (64-75) |
| Байдарочники, 26 (25-27) лет, n = 4 | 181 см (179-186) 74.0 кг (71 .0-79.0) | Р Н | 58.4 ± 3.8 (48.2-65.9) 61.4 ± 6.2 (45 .0-72.4) | 7.9 ± 0.6 (7.1-9.2) 5.8 ± 0.9 (3.3-8.0) | 25.0 ± 6.8 (19 .0-42 .0) 22.2 ±4.7 (11.3-31.5) | 54.9 ± 2.0 (51-60) 56.8 ± 0.8 (55-58) |
| Бегуны, 23 (19-33) лет, n = 8 | 177 см (168-185) 69.5 кг (59.1-80.8)
| Р Н |
58.9 ±.3.7 (52.7-70.0)
| 4,2**
6.4 ± 0.5 (4.3-8.2)
|
20.1 ± 2.5 (10.3-26.5)
|
72.4 ± 1.9 (61-80) |
| Пловцы, 21 (18-23) лет, n = 5 | 181 см (175-189) 78.3 кг (70.0-78.6) | Р Н | 74.3 ± 5.7 (65.9-85.3) 57.7 * 9.3 (37.2-79.7) | 8.6 ± 0.7 (6.2-9.9) 7.6 ± 0.5 (5.5-8.5) | 22.7 ± 0.4 (21 .0-23.2) 29.3 ± 0.4 (28.0-29.7) |
79.9 ± 1.0 (69-74) |
| Пауэрлифтеры, 25 (23-29) лет, n = 4 | 171 см (159-186) 81.3 кг (52 .0-107.0) | Р Н | 52.6 ±7.7 (42.6-66.7) 46.1±10.5 (25.3-59.7) | 2.6 ± 0.4 (2.0-3.2) 3.0 ± 0.3 (2.5-3.7) | 21.1 ± 4.8 (12.1-29 .0) 24.7 ± 1.7 (20.8-29.4) | 33.5 ± 3.06 (27-40) 40.1 ± 6.4 (32-55) |
| Ориентирщики, 52 (47-58) лет, n = 11 | 176 см (161-184) 72.7 кг (59 .5-88.0) | Р Н | 63.1 ±5.1 (31 .0-98.2) 68.8 ± 5.2 (46.5-95.7) | 4.1 ± 0.5 (1.8-6.6) 5.7 ± 0.3 (3.9-7.9) |
50.7 ±2.2 (37-62) | |
Представлено: среднее арифметическое ± ошибка среднего арифметического. В скобках показаны самые высокие и самые низкие значения.
* Эта группа состоит из спортсменов, соревнующихся в нескольких различных видах спорта.
** Среднее по трем значениям.
Максимальное потребление кислорода (МПК) во время работы ног было определено для всех субъектов либо во время работы на беговой дорожке или кручением педалей велосипеда. В некоторых группах, в которых спортивная специализация включала в себя широкое использование рук, МПК как также было измерено при проворачивании специально предназначенного велосипеда для рук и ладоней (25). Выдохнутый воздух был собран в мешки Дугласа и его объем был определен с помощью мокрого спирометра. Анализ газа был выполнен с помощью техники Holdein.
Площадь мышечных волокон у некоторых людей была определена планиметрией с фотографий образцов, используемых для определения ДПН-диафоразы. Случайно выбранные клетки, в количестве 40, в том числе 20 из каждого типа волокна, были измерены для каждого человека. Диаметры клеток были рассчитаны на основе измерения площади мышечных волокон.
РЕЗУЛЬТАТЫ
Целью этого исследования было изучение людей разных возрастов и состояний физической подготовки, некоторые из которых принимали участие в различных спортивных мероприятиях. Эффективность тематической подборки в соответствующие критерии пригодности проиллюстрировано уровнем МПК во время работы ног (таблица 1), который варьировал со средним значением от 41 (неподготовленные группы и пауэрлифтеры) до 75 (у бегунов) мл / кг мин—¹. Самая низкая величина отдельного значения составила 27 и самая высокая 80 мл / кг мин—¹. Эти значения аналогичны тем, которые ранее сообщены на основе данных от малоподвижных людей до чемпионов (22). Ранее было показано, что во время МПК работа рук достигает только 70% от работы ног (1, 25). Для групп обследованных людей в нашем исследовании эти значения были 79 и 97% для тяжелоатлетов и байдарочников, соответственно. Это иллюстрирует важность количества массы мышц, включенных в работу; и тренировку конкретных групп мышц.
Два типа волокон были определены в образцах мышц и рассмотрены в данном исследовании (рис. 1). Один обладает высокой, другой – низкой активностью АТФазы миозина при щелочном рН (9,4).
В соответствии с Barnard соавт. (2), высокая активность АТФазы миозина в условиях нашего анализа происходила в МВ с быстрыми характеристиками сокращения, низкая активность – в мышечных волокнах с медленными характеристиками сокращения. У нас, таким образом, обозначены два типа волокон, как быстро сокращающиеся (FT тип) и медленно сокращающееся (ST тип). Как показал альфа-глицерофосфат дегидрогеназы, МВ FT типа всегда имеют гликолитическую емкость выше, чем ST типа волокна. МВ FT типа, как правило, имеют более низкую окислительную способность, чем МВ ST типа. Это видно из активности ДПН-диафоразы, окислительная способность которой существует в обоих типах волокон. В центре этой сплошной среды существует область, где окислительная способность двух типов волокон совпадает. Как будет показано впоследствии, с тренировкой, окислительная способность обоих типов волокон повышается. Таким образом, любую классификацию системы на основе частичной или исключительно на окислительной способности, будет трудно объективно применить ко всем образцам мышц.
Третий тип волокна, имеющий высокую активность миозина АТФазы при щелочном рН и высокую окислительную способность, был идентифицирован в скелетных мышцах различных видов (2). Этот тип волокна был назван красным (2), С (24), быстросокращающимся (5), и αβ (10). Мы не видели этот тип волокна в изученных образцах мышц. Оgata и Murata (19), однако, выявили третий тип волокна в человеческой межреберной мышце на основе SDH и окрашивания черным суданом. Эти исследователи, однако, не рассмотрели сократительных характеристик, которые показывает АТФаза миозина.

Среднее распределение волокон в образцах дельтовидной мышцы было 46% ST типа и 45% ST типа (% быстросокращающиеся волокон = 100% — медленно сокращающихся волокон) для нетренируемых молодых и среднего возраста групп, соответственно (таблица 1). Эти образцы содержали от 14 до 60% МВ ST типа у молодых спортсменов, и от 34 до 58% МВ ST типа в средней возрастной группе. Среднее распределение волокон в боковой части латеральной широкой мышцы бедра было сравнимо с дельтовидной у молодых людей, имеющих 36% МВ ST типа и у группы среднего возраста — 44% МВ ST типа. Диапазон численности волокон этой мышцы был таким же, как и у дельтовидной, 13-73% МВ ST типа, как для молодых людей, так и для группы среднего возраста. Edström и Nyström (9) отметили обширное распространение типов волокон в скелетных мышцах человека.
Средний процент МВ ST типа в мышцах людей, которые принимали участие в тренировке на выносливость, был выше, чем в малоподвижных группах. Хотя были некоторые исключения, самый высокий процент МВ ST типа был найден в мышцах, которые были весьма активно задействованы в работе на выносливость. Среднее значение для мышц ног было около 60% МВ ST типа. Диапазон обоих МВ ST и FT типа в любой конкретной группе спортсменов тренировавшей выносливость, был похож на тот, что и у нетренирующихся групп. Тем не менее, низкие и высокие значения были выше, чем у малоподвижных групп. МВ ST и FT типа примерно поровну распределены в дельтовидной и широкой латеральной мышцы тяжелоатлетов.
Диаметры волокон и области и относительной площади, занятые каждым типом волокна в мышце были определены на 25 образцах, которые включали 15 от ног и 10 от рук (Таблица 2). Эти образцы представляли все группы и включали шесть людей, у которых были сделаны измерения, как на руках, так и ногах. Средняя площадь МВ ST типа для всех образцов, была 5423,6 и 5406,5 мкм² для широкой латеральной и дельтовидной, соответственно. Площадь МВ FT типа была на 20% больше, чем площадь МВ ST типа в обеих: латеральной широкой мышце бедра (6379,8 мкм²) и дельтовидной мышце (6295,7 мкм²). Когда сравнения были сделаны для людей, у которых имелись данные, полученные из мышц ног и рук одного человека, площади МВ ST типа были 5771,8 мкм² и 5295,7 мкм², соответственно. У этих людей, МВ ST типа ноги были на 9% больше, а МВ FT типа на 8% больше, чем аналогичные на руке. Отмеченные изменения действительно существуют между различными мышцами у одного того же человека или между людьми. Для нескольких людей (D.M., B.L,. и D.F.) МВ ST типа были более крупными, чем МВ FT типа (таблица 2, рис. 1С).
С несколькими исключениями, мышечные волокна ST и FT типа у тренированных людей были больше, чем у нетренирующихся. Тип тренировки также, как оказалось, оказывает влияние на относительный размер двух типов волокон. У спортсмена М.Н. (тяжелоатлет) волокна ноги были только на 9% больше, чем те, что на руке. Однако, как в руке, так и в ноге МВ FT типа были более чем на 60% больше, чем МВ ST типа. Напротив, МВ ST и FT типа руки байдарочника B.L. были на 26% и 64% больше, соответственно, чем МВ ноги. У спортсмена S.H., пловца, МВ ST типа руки были на 24% больше, чем МВ ноги, в то время как МВ типа FT руки на 5% меньше, чем МВ ноги. У этого испытуемого, типы волокон руки были примерно равны по размеру, но МВ ноги FT типа на 40% больше, чем МВ ST типа. Эти примеры иллюстрируют влияние хорошей подготовки рук на размер волокна. У испытуемого J.R., велосипедиста, МВ ST и FT типа мышц руки на 37% и 26% меньше, чем соответствующие МВ ноги. МВ FT типа руки были на 34% больше, чем МВ ST типа, но для ноги эта разница была только 15%. Во всех из этих примерах, площадь волокон тренированных спортсменов была больше, чем у нетренированных лиц.
Относительная площадь мышцы, занятая определенным типом волокна была совершенно иной у обычных спортсменов, по сравнению со спортсменами, занятыми тренировками на выносливость. У нетренированных людей площадь МВ ST типа колебалась от 24,5 до 53,2% от общей площади. Низкие значения спортсменов, незанятых в тренировках на выносливость включают 17,3 % для ног у тяжелоатлетов и 21,9% для ног у спринтеров. Наоборот, площадь, занимаемая МВ ST типа была 80,1% для ног бегуна на длинные дистанции (D.A.), 84,4% и 73,7% для рук и ног пловца (S.H.), соответственно, и 74,5% для рук байдарочника (B.L.).
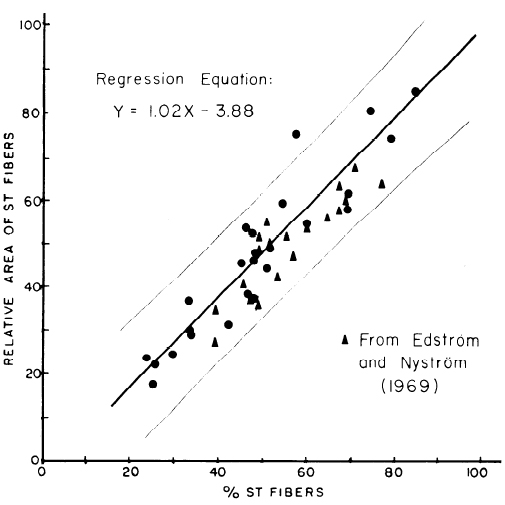
Из приведенных выше данных и таблицы 2, видно, что для большинства людей МВ FT типа больше по размерам, чем МВ ST типа. Можно также отметить, что у некоторых тренирующихся спортсменов преимущественно увеличиваются мышечные волокна обоих типов. В этих условиях распределение двух типов волокон в мышце не может свидетельствовать об относительной площади, занимаемой определенным типом волокна. Процентное соотношение между МВ ST типа и относительной занимаемой ими площади, в образце мышцы, было определено с помощью всех имеющихся данных в настоящем исследовании. Как показано на рис. 2, существует линейная зависимость между процентным распределением МВ ST типа и относительной площадью, занимаемой МВ ST типа. Это было верно даже, если образцы содержали значительные отличия в соотношении численности волокон и размерах. В данных, ранее опубликованных Edström и Nyström (9) было приведено процентное распределение и относительная площадь двух типов волокон из нескольких различных мышц мужчин и женщин. Эти данные также отображены на рис. 2. Близкие совпадения существуют между результатами настоящих исследований, и теми, что сообщили Edström и Nyström (9). Все значения обоих исследований, кроме одного несоответствия, входят в 95% доверительного интервала линии регрессии. Таким образом, процентный состав мышцы с точки зрения типов волокон свидетельствует о том, что относительная площадь каждого типа волокна вносит свой вклад в площадь всей мышцы.
Активность СДГ у нетренированных людей, как молодых и среднего возраста составляла около 3,5 и 4,4 µмоль /г мин-¹ для мышц рук и ног, соответственно (таблица 1). Активность СДГ рук и ног мышц тяжелоатлетов была на 30% меньше, чем у нетренированных людей. Активность СДГ рук и ног мышц остальных тренируемых была на 20 — 150% выше, чем малоподвижных групп. Самая высокая индивидуальная активность СДГ была найдена в мышце ноги велосипедиста (12,4 µмоль /г мин-¹) и мышцах рук пловца (9,9 µмоль / г мин-¹).
Активность СДГ была самой высокой в наиболее активных мышцах, как для подготовленных, так и для неподготовленных испытуемых. В неподготовленной группе активность СДГ мышцы ноги была на 25% больше чем мышцы руки. В тех группах спортсменов, у которых в тренировку были вовлечены ноги, активность СДГ дельтовидной мышцы была в среднем на 15-20% больше, чем у нетренированных людей. У этих же испытуемых, активность СДГ в мышцах ног была на 55 % выше, чем у рук.
Таблица 2 Размеры волокна, численность, и отношение к мышечной площади у нескольких испытуемых
| Исследуемый | Образец (рука или нога) | Группа | Диаметр волокна, мкм | Площадь, мкм² | % МВ ST типа | % площади МВ ST типа | ||
| МВ ST типа | МВ FT типа | МВ ST типа | МВ FT типа | |||||
| P.G. | Н | Нетренируемый | 75,2±2,9 (47,1-96,1) | 85,8 ± 2,0 (69,3-101,7) | 4567,5 ± 343,2 (1740-7250) | 5843,0 ± 273,9 (3770-8120) | 34,0 | 28,7 |
| C.S. | Н | Нетренируемый | 80,3±3,3 (54,4-112,2) | 93,2 ± 2,2 (71,9-l10,4) | 5234,5 ± 440,3 (2320-9860) | 6902,0 ±329,9 (4060-9570) | 30,0 | 24,5 |
| MKS | Н | Нетренируемый | 63,4 ± 2,4 (47,1-79,4) | 67.7 ±2,4 (47,1-86,0) | 3057,5 ± 273,6 (1740-4930) | 3683,0 ±250,0 (1740-5800) | 34,0 | 30,0 |
| GK | Р | Нетренируемый | 63,6 ± 2,0 (38,0-76,0) | 67,3 ±1,7 (53,8-79,2) | 3234,0 ± 181 ,2 (1135-4544) | 3594,0 ± 177,8 (2270-4918) | 48,3 | 45,7 |
| Н | 72,2 ± 3,5 (43,9-98,2) | 75,0 ± 1,4 (65,9-87,8) | 4275,0 ± 382,1 (1513-7566) | 4445,0 ± 169,1 (3405- 6053 ) | 48,6 | 47,6 | ||
| NP* | Н | Спринтер
| 79,5 ± 2,6 (54,4-98,0) | 89,4 ±2,1 (74,4-101,7) | 5060,5 ± 314,0 (2320-7540) | 6336,5 ±284,2 (4350-8120) | 26,0 | 21,9 |
| DM* | Н | Бегун
| 67,1 ±1,7 (54,4-83,3) | 58,0 ±: 1,3 (47,1-69,3) | 3581,l ± 186,2 (2320-5510) | 2668,0 ± 1,220 (1740-3770) | 75,0 | 80,1 |
| DS | Н | Бегун
| 85,1 ± 3,5 (57,7-105,3) | 105,2 ± 2,5 (81,5-124,6) | 5858,0 ± 445,8 (2610-8700) | 8776,l ± 403,2 (5220-12179) | 70,0 | 60,9 |
| DF* | Н | Бегун -средневик | 95,5 ± 3,2 (74,4-121,6) | 87,9 ± 3,4 (47,1-115,3) | 7307 8 ± 499, (4350-11599) | 6,235 ,0 ± 448,2 (1740-10439) | 55,0 | 58,9 |
| RP | Н | Бегун -средневик | 59,2 ± 2,7 (27,2-79,2) | 71 ,6 ±2,5 (50,9-98,0) | 2856,5 ± 231,8 (580-4830) | 4118,0 ± 295,9 (2030-7540 ) | 47,0 | 38,1 |
| BA* | Н | Бывший тяжелоатлет | 107,1 ± 3,8 (83,8-135,9) | 108,9 ± 3,4 (92,2-160,8) | 9,199,l ± 656,7 (5510-14499) | 9,482,9 ± 666,9 (6700-20 299) | 24,0 | 23,5 |
| MH | Н | Пауэрлифтер | 85,6 ±4,9 (47,1-113,7) | 110,8 ± 3,0 (86,0-135,9) | $035,6 ±629 (1740-10149 | 9758,1 ± 516,6 (5800-14499) | 25,3 | 23 , 5 |
| Р | 83,5±2,4 (60,8- 98,0) | 105,0± 3,3 (74,4-135,9) | 5553,5 ±303 (2900-7540) | 8917,2 ± 543,1 (4350- 14499) | 48,4 | 36,9 | ||
| JR* | Р | Велоспедист
| 83,1 ± 1,4 (71,9-96,1) | 96,2 ± 1 ,9 (76,9-108,7) | 5467,0 ± 187 (4060- 7250) | 7337,0 ± 273,7 (4640-9280) | 52,1 | 48,6 |
| Н | 104,6 ± 2,3 (86,0-127,5) | 112,2 ±2,2 (98,0-137,2) | 8,651 ,5 ± 763,9 (5800-12759) | 9,946,6 -±401 5 (7540-14789) | 51,3 | 44 , 0 | ||
| BL | Р | Байдарочник | 101,9 ± 2,6 (86,3-l29,5) | 102 ,9 ± 2,3 (88,6-118,2) | 8244 ± 570,3 (5850-13162) | 8391 ,0 ± 361,9 (5484-10968) | 57,9 | 74 ,6 |
| Н | 90,5 ± 2,7 (68,2-107,9) | 80,3 ± 1,5 (68,2-91,6) | 6544,0 ±387,5 (3656-9140) | 5100,0 ± 190,1 (3656- 6581) | 69,9 | 57,5 | ||
| SH | Р | Пловец | 88,0 ± 1,7 (71,6-101,2) | 91 ,0 ± 2,4 (77,8-105,7) | 6124,0 ± 233,9 (4022-8043) | 6552,0 ± 263,2 (4753-8774) | 85,3 | 84,4 |
| Н | 79,0 ± 2,0 (61,9- 91,6) | 93,6±1,9 (74,8-105,7) | 49540 ± 2,37 6 (2925- 6,581) | 6928,0 ± 266 ,9) (4387-8774 | 79,7 | 73 ,7 | ||
Данные означают: среднее арифметическое ± ошибка среднего арифметического Значения в скобках максимальные и минимальные *См. таблицу 4 для доп. характеристик
В группе ориентировщиков среднего возраста активность (СДГ) латеральной широкой была на 40% выше, чем дельтовидной мышцы. Еще большие различия существовали между неподготовленными группами и теми испытуемыми, в чьих спортивных мероприятиях имело место широкое использование мышц ног или рук. Различия между мышцами у одних и тех же испытуемых стали более выраженным в хорошо подготовленных группах. В случае велосипедистов, активность СДГ мышцы ноги была в 2,5 раза больше, чем у малоподвижных людей. Деятельность рук этих субъектов была на 70% выше, чем у нетренированных, но только на 55% для ног. Деятельность СДГ дельтовидной мышцы у байдарочников и пловцов была в 2,2 и в 2,4 раза выше, соответственно, чем у неподготовленных групп. У этих испытуемых активность СДГ была выше в мышце руки, чем ноги. Эти результаты свидетельствуют о специфике локализации тренировочного эффекта.
Хотя активность ФФК не была определена для всех групп, не было никаких различий между группами или между мышцами в пределах данной группы. Средние значения были от 19 до 29 µмоль /г мин—¹, самое низкое значение найдено в группе субъектов, занятых в различных спортивных мероприятиях, а самая высокая величина найдена в мышцах ног пловца (Таблица 1).
Значение активности фермента и композиции волокна исследовали также с помощью биопсии латеральной широкой мышцы, взятых образцов из различных мест у 18 испытуемых (в том числе тренируемых и нетренируемых). Место отбора проб в этих экспериментах было на 12-16 см выше коленной чашечки на глубине 4 см в мышце. Альтернативные места для взятия проб были 4 см выше или ниже этой точки. Стандартное отклонение различий между этими образцами составило 0,46 и 3,86 µмоль /г мин—¹, для активности СДГ и ФФК, соответственно, и 4,6 для процентного распределения волокон. Хотя это представляет собой небольшую степень изменчивости, оно иллюстрирует необходимость стандартной выборки мест для проб. Вариация в глубине погружения иглы для биопсии также может повлиять на активность фермента, размер и распределение волокон. Тем не менее, это не было определено в настоящем исследовании.
РНК в мышцах дельтовидной и латеральной широкой определялась только у велосипедистов, тяжелоатлетов, ориентировщиков среднего возраста и пловцов. Средние значения для этих групп были 1,5 и 1,7 µг /г для руки и ноги, соответственно. Не выявлены существенные различия между группами или между двумя мышцами исследуемых. Поскольку эти группы включают широкий диапазон возрастов (17-58 лет) и различные виды спорта, кажется, что лишь незначительные различия могут быть объяснены этими факторами.
Концентрация мышечного гликогена у тренированных людей, как правило, выше, чем у малоподвижных (таблица 3). Было также установлено, что для одного субъекта содержание гликогена было самым высоким в мышцах, которые были наиболее задействованы в спортивной деятельности. Примером является испытуемый J.R .(велосипедист), у него высокое содержание гликогена было обнаружено в мышцах ног, обратный результат получен у S.H. и B.L. (пловец и байдарочник). На основе окрашивания PAS, кажется, что не существует единого шаблона для содержания гликогена в двух типах волокон в скелетных мышцах. Примеры из некоторых образцов, наблюдаемых в этом исследовании, представлены на рис. 3. В некоторых случаях было невозможно выявить различия в концентрации гликогена между двумя типами волокон (рис. 3, А и С). Содержание гликогена в ткани на рис. 3,4 было низким (84 mм единиц глюкозы/кг), а на рис. 3C достаточно высоким (178 mм единиц глюкозы/кг). В других примерах (рис. 3, Е и G), образцы варьируют: наиболее интенсивно окрашиваются то МВ ST то МВ FT типа. Во многих мышцах образцы чередовались (рис. 3G). Способность МВ ST типа накапливать гликоген иллюстрируется испытуемым D.M., у которого мышцы содержат 75% МВ ST типа и высокое содержание гликогена (127 mм единиц глюкозы/кг).
ОБСУЖДЕНИЕ
Идентификация только двух типов волокон в мышцах человека в настоящем исследовании согласуется с несколькими предыдущими докладами (7, 9, 16). В отличие от предыдущих наблюдений мы имеем выбранную идентификацию волокон на основе сократительных характеристик. Высокая активность СДГ в мышцах тренируемой группы, соответствует выводам, сообщенных Varnauskas и соавт. (26), Morgan и др. (15). Активность DPSH-диафоразы в МВ ST и FT типа у тренированных людей была выше, чем у малоподвижных (рис. 1). Это свидетельствует о адаптируемости окислительной способности в обоих типах волокна. В некоторых тренируемых группах (рис. 1), активность DPSH-диафоразы МВ FT типа была больше, чем в МВ ST типа в мышцах нетренированных групп. Мышечные волокна ST типа в мышцах высококвалифицированных спортсменов были более окислительными чем МВ ST типа у нетренированных. Это характеризовалось темным субсарколемным окрашиванием DPSH-диафоразы. У спортсменов значение самой высокой активности СДГ DPSH-диафоразы МВ FT типа приближалось к значению МВ ST типа.
Таблица 3. Концентрация гликогена в мышце руки и ноги нетренируемых и представителей тренируемых групп
| Испытуемый | Вид спорта | Гликоген, мМ единиц глюкозы / кг | |
| Нетренируемые | 93,3* (75-120) | 80,7** (53-111) | |
| J.R. | Велосипедист | 90 | 153 |
| S.H. | Пловец | 223,0 | 122 |
| B.L. | Байдарочник | 105 | 79 |
| D.M. | Бегун | 127 | |
| D.S. | Бегун | 173 | |
| D.F. | Бегун на средние дистанции | 178 | |
В скобках показаны границы наблюдений (самое высокое и самое низкое)
* Среднее по трем людям. **Среднее значение для 10 испытуемых.
Возникает вопрос, может ли существующее процентное распределение определенного типа волокна в скелетной мышце быть изменено с помощью тренировки. Morgan и соавт. (15) сообщили, что у человека процент красных волокон возрастает после тренировки. Barnard и его коллеги (3) также отметили, что процент красных волокон в мышце морской свинки увеличивается после тренировки. Следует отметить, что преобразование типов волокон, как сообщает Barnard c соавт. не требует изменений сократительных свойств. Кроме того, Morgan и др. определили волокна как красные или белые только на основе активности СДГ. Поскольку тренировка приводила к изменению активности СДГ (3, 11), и как было показано в настоящем исследовании, окислительная способность МВ FT типа может резко меняться, то будет легко идентифицировать МВ ST и FT типа после тренировки от одного гистохимического определения активности СДГ. Однако, Guth и Yellin (10) отметили постепенное уменьшение (высокой активности) АТФазы миозина окрашенных волокон у камбаловидной крысы при компенсаторной гипертрофии. Хотя на вопрос указанный выше, невозможно ответить с помощью данных этого исследования, мы не можем исключить возможность того, что такое преобразование волокон может происходить с помощью тренировок. Широкий диапазон композиций волокон в различных группах может быть причиной такой конверсии волокон из одного типа в другой. С другой стороны, некоторые из видов спорта на выносливость действительно давали относительно высокий процент МВ ST типа у испытуемых. Это может быть связано с выбором этого вида спорта спортсменом, обладающим природными способностями. Необходимы дополнительные исследования, чтобы уточнить этот момент.

Существовали немалые различия в составе мышечных волокон и активности ферментов, и исполнительными характеристиками некоторых из лучших спортсменов, изученных в данном исследовании. Несколько примеров этого представлены в таблице 4. Сравнив показатели испытуемых J.R .и R.P. с D.M., видно, что высокая активность СДГ и МПК схожа, но сильно отличается состав мышечных волокон. Испытуемый R.P., чемпион мира, байдарочник, J.R., велосипедист, у обоих было примерно равное распределение этих двух типов волокон в ноге и мышце руки, в то время как у D.M., бегуна на длинные дистанции, было преобладающим количество МВ ST типа (75%) в латеральной широкой мышце. Высокие уровни активности СДГ в мышцах J.R. и R.P. были результатом высокой окислительной способности в обоих типах волокон, что повлияло на активность DPSH-диафоразы. Еще одно различие между этими спортсменами — их способность выполнять скоростную работу. Оба испытуемых R.P. и J.R. обладали высокой способностью к спринтерской работе. R.P. имеет лучшие результаты в беге на короткие дистанции, в то время как испытуемый D.M. показывает лучшие результаты на более длинных дистанциях и имеет ограниченные спринтерские способности. J.R. мог выполнять длительную работу на выносливость, а затем спринт в конце соревнования. D.F., с 55% МВ ST типа, является бегуном на средние дистанции (4:04 = миля и 1:51 =полмили). N.P. (74% МВ FT типа) является спринтером (9,3 с 100 ярдов). Различия в композиции мышц может быть причиной, по крайней мере, частично, для этих характеристик. Интересно также отметить, что в МПК испытуемого R.P. было выше в работе руки, чем ноги. Это было, несомненно, связано с высокой тренировкой мышц рук байдарочника. Подобные сравнения также могут быть сделаны между A.B., пловцом, который соревновался в плавании на 400-1500м и испытуемыми J.R. и R.P.
Таблица 4. Композиция мышечных волокон, активность ферментов, МПК, и исполнительные характеристики нескольких испытуемых
| Испытуемый, возраст, вес, рост | Конечность для проб (рука или нога) | % МВ ST типа | Активность ферментов µмоль /г мин-¹ | МПК л/мин | Комментарии | |
| СДГ | ФФК | |||||
| J.R. 33 года, 175 см, 75 кг | Р | 51.3 | 6.73 | 19 36 | Велосипедист, 5-ти кратный чемпион Швеции | |
| Н | 52.1 | 12.38 | 21 .37 | 5,50 | ||
| C.S. 26лет, 189 см, 74 кг | Р | 39.5 | 5.49 | 4.80 | Велосипедист троекратный чемпион мира, 1969; 5-ти кратный чемпион Швеции | |
| Н | 72.5 | 8.21 | ||||
| R.P. 27лет, 186 см, 79 кг | Р | 48.2 | 9.24 | 22.98 | 4.77 | Байдарочник; олимпийская золотая медаль, 1964; олимпийский серебряная медаль, 1968; пятикратный чемпион мира, последний раз в 1971 |
| Н | 45.0 | 5.65 | 31.45 | 4.55 | ||
| A.B. 19лет, 175 см, 70 кг | Р | 71 .8 | 9.60 | 20.97 | Пловец; 400 м. — 4: 05.8; 1500м — 16:33.4 | |
| Н | 66.4 | 8.06 | 28.83 | 5.20 | ||
| D.M. 20лет, 170 см, 62 кг | Р | Бегун; 4-й в NCAA Чемпионате по кроссу, 1971; 28:15 при пробегании 6 миль | ||||
| Н | 75,0 | 8.03 | 15.07 | 4.95 | ||
| D.F. 21лет, 178 см, 69 кг | Р | Бегун на средние дистанции; 1:51-880 yd; 4:04 — миля | ||||
| Н | 55.0 | 5.14 | 26.53 | 4.78 | ||
| NP -22лет, 165 см, 61 кг | Р | Спринтер; 9.3 100 yd | ||||
| Н | 26.0 | 3. 95 | 28.34 | 3.59 | ||
| B.A. 31год, 191 см, 87 кг | Р | Бывший тяжелоатлет вес; деятельность в спортивной рекреации | ||||
| Н | 24.0 | 4.67 | 23.12 | 4.39 | ||
Еще один интересный пример – C.S., который в 1969 году имел МПК 5,6 л/мин и занял третье место на чемпионате мира по велоспорту. В связи с неспособностью продолжать свои тренировки МПК отказалось 4,8 л/мин на момент исследования. Тем не менее, его широкая латеральная мышца бедра содержит 73% МВ ST типа и имеет активность СДГ 8,2 µмоль/гмин—¹. Эти характеристики свидетельствуют о высокой производительности аэробной работы у B.A., бывшего тяжелоатлета, который по-прежнему занимается спортом, МВ FT типа ноги преобладали (76%), в то время как активность СДГ была средней (4,47 µмоль/г мин—¹).
Установлено, что в мышцах некоторых нетренирующихся субъектов, процентное содержание МВ ST типа может быть равно 70-85%, , что соответствует показателям спортсменов, тренирующихся на выносливость, хотя МПК и активность СДГ и DPSH-диафоразы были намного ниже. В каждом случае высококвалифицированные спортсмены на выносливость обладали высокой активностью СДГ и DPSH-диафоразы. Эти две характеристики, как представляется, хорошо связаны между собой. Если активность СДГ и DPSH-диафоразы может резко меняться в обоих типах волокон, то спортсмены с высоким процентом МВ FT типа еще могут обладать чрезвычайно высокой окислительной способностью, приспособляемость волокон для аэробного метаболизма может быть более важным фактором в тренировке и исполнительности, чем основной состав мышцы.
Один вопрос в настоящее время все еще открыт. Что является лимитирующим фактором МПК: окислительная способность мышц или пропускная способность сердечно-сосудистой системы? К этому вопросу можно подходить с точки зрения окислительной способности мышц на основе активности СДГ. Для такого подхода, следует отметить, что активность СДГ была измерена при 25 С в настоящем исследовании, и что при нормальной температуре тела 37 С происходит 2,2-кратное увеличение активности (6). Если предположить, что объем потребляемого кислорода скелетных мышц равен скорости конверсии сукцината на 1 кг скелетных мышц, широкая латеральная мышца бедра у испытуемого малоподвижной группы может потреблять 0,6 л кислорода/мин. Для велосипедиста (активность СДГ 11,0 µмоль/г мин—¹), это значение будет 1,6 л кислорода/кг мин. В таких условиях, потребление кислорода всем телом во время максимальной работы может быть равным 6-7 кг для мышц человека из нетренируемой группы и от 3-3,5 кг для мышц велосипедистов. Это позволяет предположить, что как для малоподвижных, так и для тренируемых испытуемых, окислительная возможность мышечной массы, используемая в упражнении для ног, будет превышать известные значения МПК. Эти расчеты основаны на предположении, что все компоненты окислительного метаболизма функционируют со скоростью реакции СДГ, что маловероятно. Однако, это было продемонстрировано (12), большинство компонентов из цикла Кребса и системы транспортировки электронов увеличивается пропорционально друг другу в процессе тренировки. Тем не менее, активность СДГ представляет все митохондриальные белки. Исходя из этих соображений и активности СДГ в этом исследовании, окислительная способность мышц ноги велосипедиста в 2,5 раза больше, чем у малоподвижного испытуемого. МПК высококвалифицированных спортсменов, однако, не было в 2,5 раза больше, чем у нетренируемых испытуемых. Эти рассуждения, кажется, указывают на сердечно-сосудистую систему, как на устанавливающую верхний предел МПК.
Что тогда является важным критерием большой аэробной способности скелетных мышц спортсменов? Очевидно именно увеличение окислительной способности МВ FT типа, и как результат, способность этих волокон более эффективно использовать запасы гликогена в аэробных условиях и окислять жирные кислоты. Таким образом, наступление утомления этих волокон будет отложено. Это увеличение окислительной возможности МВ FT типа также может способствовать снижению уровня лактата во время субмаксимальной работы, которая происходит после тренировки.
Как указано выше, кажется маловероятным, что способность тела использовать кислород во время максимальной работы ограничена окислительной способностью мышечной клетки. Несмотря на это, наблюдались глубокие различия между мышцами тренируемой и нетренируемой группы людей. Выносливость спортсмена была охарактеризована высоким процентом МВ ST типа в его тренированных мышцах. Окислительная способность этих волокон и МВ FT типа была значительно выше, чем в его менее активных мышцах или в мышцах нетренированных людей. Степень, с которой регулярная тренировка способствовала этой наблюдаемой разнице, не может быть определена путем настоящего исследования. Наблюдаемые результаты показывают, что физическая подготовка человека не влияет на состав волокон, однако влияет на окислительную способность отдельных мышечных волокон.
Это исследование было частично поддержано Исследовательским Советом Федерации Спорта Швеции и Советом Швеции по Медицинским Исследованиям (14X-2203). B. Armstrong и C. W. Saubert IV являются кандидатами Национального Научного Фонда.
REFERENSES:
- ASMUSSEN, E., AND I. HEMMINGSEN. Determination of maximum working capacity at different ages in work with the legs or with the arms. Stand. J. Clin. Lab. Invest. 10: 67-71, 1958.
- BARNARD, R. J., V. R. EDGERTON, T. FURUICAWA, AND-J. B. PE’I’ER. Histochemical, biochemical and contractile properties of red, white, and intermediate fibers. Am. J. Physiol. 220: 410-414, 1971.
- BARNARD, R. J., V. R. EDGERTON, AND J. B. PE’TER. Effect of exercise on skeletal muscle. I. Biochemical and histological properties. J. Appl. Physiol. 28: 762-766, 1970.
- BERGSTROM, J. Muscle electrolytes in man. Scand. J. Clin. Lab. Invest. Suppl. 68, 1962.
- BURKE, R. E., D. N. LEVINE, F. E. ZAJAC III, P. TSAIRIS, AND W. K. ENGEL. Mammalian motor units : physiological-histochemical correlation in three types of cat gastrocnemius. Science 174 : 709-712, 1971.
- COOPERSTEIN, S. J ., .1. LAZAROW, AND N. ,J. KURWSS. A microspectrophotometric method for the determination of succinic dehydrogenase. J. Biol. Chem. 186 : 129-139, 1950.
- DUBOWITZ, B., AND , A. G. E. PEARSE. .A comparative histochemical study of oxidative enzymes and phosphorylase activity in skeletal muscle. Histochemie 2 : 105- 1 17, 1960.
- DUBOWITZ ,V. Cross-innervated mammalian skeletal muscle : histochemical, physiological and biochemical observations. J. Physiol., London 193 : 48 l-496, 1967.
- ENDSTROM L,. , AND B. NYSTROM. Histochemical types and sizes of fibres of normal human muscles. Acta Neurol. Stand. 45: 257-269, 1969
- GUTH, L., AND H. YELLIN. The dynamic nature of the so-called “fiber types” of mammalian skeletal muscle. Exptl. Neurol. 31:277-300, 1971.
- HOLLOSZY, J. 0. Biochemical adaptations in muscle. Effects of exercise on mitochondrial oxygen uptake and respiratory enzyme: activity in skeletal muscle. .J. Biol. Chem. 242 : 2278-2282, 1967.
- HOLLOSZY, J. 0., L. B. OSCAI, I. J. DON, AND P. A. MOLE. Mitochondrial citric acid cycle and related enzymes: adaptative response to exercise. Biochem. Biophys. Res. Commun. 40: 1368-1373, 1970.
- KARLSSON, J., B. DIAMAN’I’, AND B. SALTIN. Muscle metabolites during submaximal and maximal exercise in man. Scand. J. Clin. Lab. Invest. 26 : 385-394, 1971.
- MORGAN, P. A., AND J. 0. HOLLOSZY. Exercise-induced increase in the capacity of skeletal muscle to oxidize palmitate. Proc. Sot. Exptl. Biol. Med. 134: 789 — 792, 1970
- MORGAN, T. E., L. A. COBB, F. A. SHORT, R. Ross, and D. R. GUNN. Effects of long-term exercise on human muscle mitochondria. In : Muscle Metabolism During Exercise edited by B. Pernow and B. Saltin. New York: Plenum, 1971, p. 87-95.
- MORRIS, C. J. Human muscle fibre type grouping and collateral re-innervation. .J. .Neurol. Neurosurg . Psychiat. 32 : 440-444, 1968
- MUNRO, H.N., AND L.FLECK. The determination of nucleic acids. Methods Biochem. Anal. 14: 113-176, 1966
- NOVIKOFF A. B., W. SHIN, AND J. DRUCICER. Mitochondrial localization of oxidation enzymes: staining results with two tetrazolium salts. J. Biophys. Biochem. Cylol. 9 : 47-61. 1961
- OGATA, T., AND F. MURA’I’A. Cytological features of three fiber types in human striated Muscle. Tohoku J. Exptl. Med. 99: 225-245. 1969.
- PADYKULA, H. A., AND E. HERMAN. The specificity of the histochemical method of adenosine triphosphatase. J. Histochem. Cylothem.3: 170-195, 1955
- PEARSE, A. G. E. Histochamistry — Theoretical and Applied. Boston, Mass. : Little, Brown, 1967, Appendix 9 p. 832.
- 22 SALWN, B., AND P.-O. ASTRAND. Maximal oxygen uptake in athletes. J. Appl. Physiol. 23 : 355-358, 1967
- 23 .SHONK, C. E., AND G. E. BOXER. Enzyme: patterns in human tissue. I. Methods for the determination of glycolytic enzymes. Cancer Res. 24 : 709-724, 1964.
- 24 STEIN, J . M., AND H. A PADYKULA. Histochemical classification of individual skeletal muscle fibers of the rat. Am. J. Anat. 110 : 103-115, 1962.
- SI’ENBERG, ,J ., P.-O. ASTRAND, B. EKBLOM, J. ROYCE, AND B. SALTIN. Hemodynamic response to work with different muscle groups, sitting and supine. J. Appl. Physiol. 22 : 61-70, 1967
- VARNAUSKAS, E., P.BJORNTORP, M.FAHLEN, I.PREROVSKY, AND J. STENBERG. Effects of physical training on exercise blood flow and enzymatic activity in skeletal muscle. Cardiovascular Res. 4 : 418-422, 1970.
- WATTENBERG, L.W., AND J.L. LEONG. Effects of coenzyme Q19 and menadione on succinate dehydrogenase activity as treasured by tetrazolium salt reduction. . J. Histochem. Cylochem. 8: 296-303, 1960
Перевод А.А. Крестининой, А.В. Самсоновой
1972_Gollnik-et-al_perevod.pdf